L'évolution du vivant expliquée à ma boulangère
Chapitre 3
3.1 Comment d’une espèce passer à deux espèces et qu’est-ce qui les différencie ?
Forts de ces nouvelles connaissances, nous pouvons réfléchir au vivant dans son ensemble. Comment relier les êtres vivants entre eux ? Qui est apparenté ? Qui est l’ancêtre de qui ? Comment faire pour répondre à ces question ? Si vous avez suivi ce qui précède, vous pouvez déjà répondre à la question du titre.
Tout d’abord, toutefois, définissons la notion d’espèce : d’après Ernst Mayr1, une espèce est « une communauté reproductive de populations, reproductivement isolée d’autres communautés et qui occupe une niche particulière dans la nature ». Le pilier de cette définition est l’idée d’interfécondité entre les populations. Donc, pour que deux populations, d’une espèce sexuée, appartiennent à la même espèce, il faut que les individus qui les composent puissent se croiser entre eux, et que leurs descendants le puissent également.
Ainsi, l’âne et le cheval produisent des mulets ou des bardots, selon le sens du croisement, qui sont des animaux stériles. L’âne et le cheval n’appartiennent donc pas à la même espèce. Par contre, le Saint-Bernard et le berger allemand peuvent avoir une descendance fertile sans problème : ils appartiennent à la même espèce.
Cependant, des hybrides fertiles sont parfois obtenus, en captivité, entre deux espèces sauvages différentes (notamment chez les plantes). Cela ne signifie pas qu’il faille les regrouper dans une seule et même espèce, car pour cela il faudrait que ces hybrides soient aussi produits dans le milieu naturel, ce qui n’est généralement pas le cas, pour des raisons diverses. Par exemple, elles ne fleurissent pas tout à fait à la même période, ne vivent pas au même endroit, ou n’occupent pas les mêmes « niches écologiques », c’est-à-dire qu’elles n’occupent pas tout à fait le même milieu, où n’y partagent pas les mêmes conditions de vie, ce qui entrave leur mise en contact. Néanmoins, l’obtention d’hybrides en conditions contrôlées permet de suspecter un ancêtre commun récent. Pour les chiens, qui se croisent en captivité... eh bien, leurs conditions de vie naturelles ne sont-elles pas la symbiose avec l’homme ? Par conséquent, les croisements étant toujours possibles entre différentes races, nous n’avons vraiment affaire qu’à une seule et même espèce.
Donc, imaginez une population d’une espèce donnée, qui se retrouve géographiquement isolée du reste de l’espèce. Par exemple, elle peut se trouver sur une île qui vient de se former, ou être isolée à cause d’une rivière, d’une éruption volcanique, ou d’une migration comme chez les criquets. Ces deux populations, sans contact, vont voir leurs fréquences
alléliques se modifier de façon différente, par le hasard de la dérive
génétique. En outre, elles peuvent éventuellement être soumises à des
pressions de sélection différentes. Au bout d’un certain nombre de générations, les différences accumulées
entre ces deux populations, simplement par dérive et sélection, voire éventuellement avec quelques mutations, seront tellement importantes que les individus ne pourront
plus se croiser, même si on les mélangeait de nouveau. Elles sont
isolées génétiquement : plus aucun allèle* ne peut passer d’une
population à l’autre, il n’y a plus aucun flux de gènes*.
Comment se représenter un « flux de gènes » ? Lorsque l’on s’amuse à faire de la génétique au niveau d’une population, on se représente chaque population sous forme d’un grand sac, contenant les allèles de chaque individu pour un gène donné. Pour chaque population, on a donc un sac avec une certaine proportion de chaque allèle, proportion qui se modifie au cours des générations en fonction de la sélection naturelle, de la dérive, et des mutations éventuelles.
Quand des individus migrent d’une population à l’autre (et se croisent à elle), ils véhiculent des allèles d’un sac à l’autre : c’est un « flux » d’allèles. Quand deux populations sont physiologiquement incapables de se croiser, le flux d’allèles est nul, même si les individus vivent au même endroit. On dit que les populations sont isolées reproductivement.
On distingue deux sortes d’isolement reproducteur :
| – | l’isolement pré-reproducteur : la fécondation ne peut avoir lieu, parce que les partenaires ne peuvent se rencontrer dans la nature, pour cause d’occupation de différentes niches écologique, ou bien ne s’identifient pas comme partenaires (par exemple, essayez de croiser un chien et un chat...), ou alors leur croisement est rendu impossible par un obstacle mécanique (la taille par exemple, ou la forme des organes reproducteurs), écologique (modification de la période de reproduction par exemple), ou physiologique (par exemple, le spermatozoïde n’arrive pas à trouver son chemin dans les voies génitales de la femelle, ou à pénétrer dans l’ovule). |
| – | l’isolement post-reproducteur : la fécondation a lieu, mais l’embryon ne peut se développer, ou bien il se développe, mais il est stérile, comme le mulet. |
Souvent, la mise en place de cet « isolement reproducteur » se fait graduellement. L’hybridation est possible, mais difficile, puis devient impossible. Mais parfois, des différences sur quelques gènes suffisent à la rendre impossible immédiatement.
Le phénomène d’apparition des espèces se nomme spéciation. Depuis l’apparition de la vie, les populations se séparent et se diversifient, les espèces s’éloignent les unes des autres, formant un arbre partant d’une origine unique.
Nous avons vu la spéciation consécutive à une isolement géographique de deux populations. Parfois, le phénomène ne nécessite pas d’isolement géographique. Dans une
même région, deux populations s’isolent dans des niches écologiques différentes. Par exemple, elles peuvent se mettre à utiliser des ressources alimentaires différentes. Il existe plusieurs formes de spéciation, que l’on distingue et nomme
selon la nature du phénomène qui précède l’isolement reproducteur.
Comme vous l’avez sûrement compris, la notion d’espèce est à prendre avec des pincettes, elle n’est pas absolue. D’une part, elle ne s’applique avec cette définition qu’aux sexués. D’autre part, de nombreuses espèces sont actuellement en cours de spéciation... Ce qui les rend difficile à séparer et à nommer, et entraîne d’interminables querelles de spécialistes.
Cela ne remet pas l’évolution en cause, bien au contraire. Il faut juste savoir de quoi on parle quand aborde la notion d’espèce, et ne pas l’appliquer à tort et à travers.
3.2 Pourquoi certaines espèces semblent n’avoir pas changé depuis des millions d’années ?
Si vous avez suivi, vous savez que des mutations se produisent tout le temps. Vous savez aussi que les conditions environnementales à la surface du globe sont changeantes et donc que les pressions de sélection se modifient avec le temps. Comment expliquer alors la présence des « fossiles vivants » comme le cœlacanthe ? La réponse est simple : les fossiles vivants n’existent pas. Les organismes ont changé, mais cela ne se voit pas morphologiquement. Pourtant, il y a eu des mutations dans leur génome (l’ensemble de leur information génétique), et des modifications des fréquences alléliques entre la population ancestrale et la population moderne. Leur physiologie ou leur métabolisme peuvent s’être modifiés, ils ne sont pas restés « figés » dans le temps.
Si cela vous semble clair, vous pouvez maintenant comprendre qu’il est faux de dire que l’homme descend du singe ! Eh oui, le chimpanzé, que l’on utilise couramment pour illustrer cette idée, est un animal actuel ! La phrase juste est donc : « l’homme est un singe (ou pour être plus précis, un primate) », ou encore « l’homme a un ancêtre commun avec le chimpanzé ». Et notre ancêtre commun avec le chimpanzé est très récent, alors que celui que nous avons avec des espèces qui sont encore plus différentes de nous est plus éloigné dans le temps. Par exemple, notre ancêtre commun avec le poisson rouge est vieux de plusieurs centaines de millions d’années. Et notre ancêtre avec le brin d’herbe est plus vieux encore. Et notre ancêtre avec les bactéries, il remonte aux origines de la vie ! Nous sommes tous cousins.
 |
Il est d’ailleurs intéressant de constater que tous les êtres vivants ont un même ancêtre commun. Comment le sait-on ? Parce que nous partageons tous un certain nombre de molécules, précises, très conservées. Les bactéries, les archées et les eucaryotes, qui représentent, ainsi que nous le verrons par la suite, les trois grands domaines du vivant, ont ainsi une base physiologique commune, notamment parce que nous utilisons tous de l’ADN et que nous partageons les mécanismes qui y sont liés. Il est beaucoup plus probable que ces mécanismes ne soient apparus qu’une fois, plutôt qu’ils soient apparus plusieurs fois indépendamment. Histoire de probabilités.
3.3 Comment dater les ancêtres ?
On
peut dater les fossiles, c’est une méthode efficace, fiable et qui a
fait ses preuves. Mais ce n’est pas cette méthode qui nous intéresse
aujourd’hui.
Comme
vous avez pu le voir plus haut, la spéciation se déroule toujours de la
même manière : une population ancestrale, qui produit deux
populations filles (ou plus). Comment
les différencie-t-on ? En analysant leur ADN. Comme les mutations se produisent ponctuellement au
cours du temps, et ont tendance à plus ou moins s’accumuler, en fonction de la façon dont elles sont soumises à la sélection naturelle, on peut dater le moment de la
divergence entre les deux populations ! Concrètement comment fait-on ?
Pour
cela, il faut trouver des séquences homologues, pour pouvoir les comparer. Rappelons-nous que des
séquences sont des fragments d’ADN, « écrits » dans l’alphabet à quatre lettres correspondant aux bases azotées A,T, G, C. Ainsi que nous l’avons vu précédemment, les
séquences homologues sont donc des gènes ou des fragments d’ADN qui ont
la même origine, c’est à dire qu’ils sont tous les deux dérivés de la
même séquence ancestrale. Par exemple, notre hémoglobine et celle du
chimpanzé sont homologues, car elles sont dérivées de celle de notre
ancêtre commun. En effet, c’est une molécule très complexe, partagée par tous les vertébrés : il est invraisemblable qu’elle soit apparue indépendamment chez toutes ces espèces.
Donc,
quand nous avons nos deux séquences, nous comptons le nombre de différences entre
elles. Et enfin, sachant la fréquence des mutations (par génération), cela nous donne une idée du temps qu’il a fallu pour que toutes ces différences
s’accumulent. C’est le principe général, celui de l’« horloge moléculaire » de Kimura, vu plus haut. Bon, la réalité est un peu
plus complexe... et nécessite pas mal de calculs.
Entrons un peu dans les détails, sans aborder les calculs. Bien
entendu, on ne prendra pas le même type de séquences si on veut
analyser des espèces proches que si on veut analyser des espèces
éloignées. Dans
le premier cas, on prendra des séquences neutres, c’est à dire non-codantes, non régulatrices, qui accumulent en peu
de temps beaucoup de mutations car elles ne sont pas soumises à la
sélection naturelle. Cependant, si ces séquences ont divergé depuis
longtemps, plusieurs mutations peuvent avoir eu lieu au même endroit (fig 3.1 ).

Ainsi,
le calcul risque d’être faux si les organismes ont divergé depuis trop
longtemps. Dans ce cas (par exemple des espèces qui ont divergé il y a
plusieurs centaines de millions d’années), on prendra donc des
séquences plus ou moins soumises à la sélection, car même dans ce cas, elles auront accumulé des différences avec le temps, mais beaucoup moins que les séquences neutres. Il y a notamment des molécules (comme
les ribosomes) que l’on retrouve chez tous les êtres vivants (ce qui
est un des arguments pour expliquer que TOUS les êtres vivants, les
animaux, les plantes, les champignons, les bactéries, ont le MÊME
ancêtre commun). Elles ont servi à élaborer l’Arbre du Vivant, car
elles ne sont pas très différentes entre tous les êtres vivants : on
peut donc les utiliser pour comparer des groupes d’espèces très éloignées.
Avec
cette méthode, on peut aussi parfois (pas toujours) avoir une idée de
l’aspect ou de la physiologie de l’ancêtre. Ainsi, si plusieurs espèces issues d’un ancêtre commun partagent toutes un certain
caractère, sauf une, il est très probable que ce caractère ait été
aussi porté par l’ancêtre, sinon, cela signifierait qu’une mutation
identique a eu lieu plusieurs fois, ce qui est très improbable. L’autre possibilité est que l’espèce en question soit le groupe frère de l’ensemble des autres, et que la mutation ait été introduite après les divergence. La figure 3.2 illustre ces deux cas. On choisit toujours l’hypothèse qui nécessite le moins de mutations, qui est dite la plus parcimonieuse.
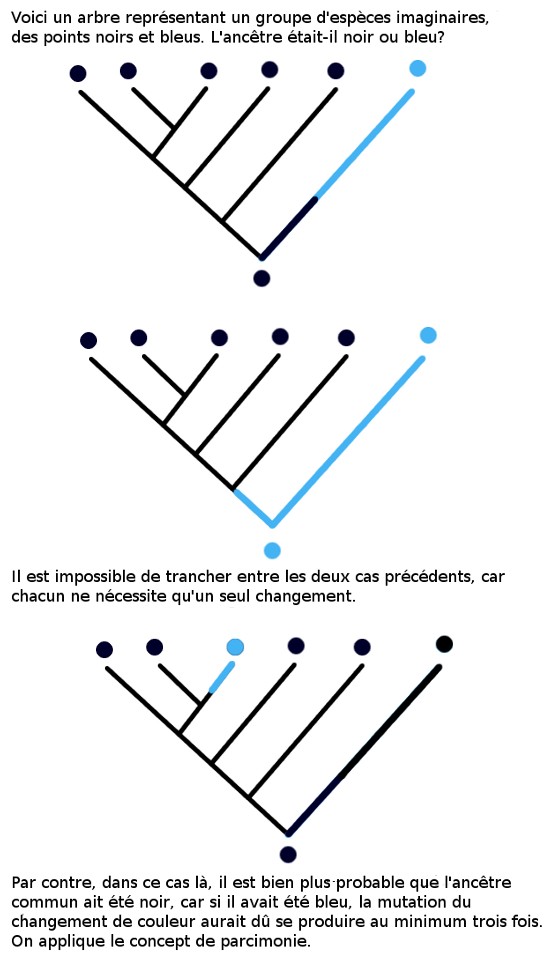
Prenons un exemple concret : l’ancêtre des mammifères avait-il un long cou ? Les girafes ont
un long cou, mais les autres mammifères ont généralement un cou assez
court. Si l’ancêtre commun des mammifères avait eu un cou long, alors
cela voudrait dire que tous les mammifères à part les girafes ont perdu
cette caractéristique. L’inverse est infiniment plus probable ! Sans
compter que les fossiles des premiers mammifères n’ont pas un cou très
long.... ce qui confirme l’hypothèse du cou ancestral court.
Nous
avons vu que les mutations étaient aléatoires. Il est aussi très rare
d’en trouver deux qui aient le même effet. Pourtant, on observe souvent dans la nature des espèces éloignées qui se ressemblent plus ou moins. Ainsi, les baleines, qui sont des mammifères, ont une forme hydrodynamique
comme les poissons. On parle alors de
convergence : des causes différentes (mutations différentes sur des
gènes différents) conduisent à des similarités morphologiques.
Cela
peut conduire à une erreur d’interprétation si on se limite à la
morphologie, mais l’analyse génétique permettra de rectifier le tir, en
constatant que les espèces que l’on croyait proches (baleine et requin
par exemple) sont en réalité très différentes.
3.4 À quoi ressemble-t-il, cet arbre ?
Dans un premier temps, il est important de comprendre qu’au fur et à mesure des découvertes, les scientifiques modifient en conséquence la structure de l’arbre du vivant, qui n’est pas parfait. Il change donc un peu d’année
en année, et est probablement amené à se modifier encore dans le futur.
Toutefois, chaque étape nous rapproche un peu plus d’une représentation correcte de la réalité.
Maintenant, voyons à quoi ressemble l’arbre tel qu’il est représenté d’après les données actuelles2. Examinons la figure 3.3 3.
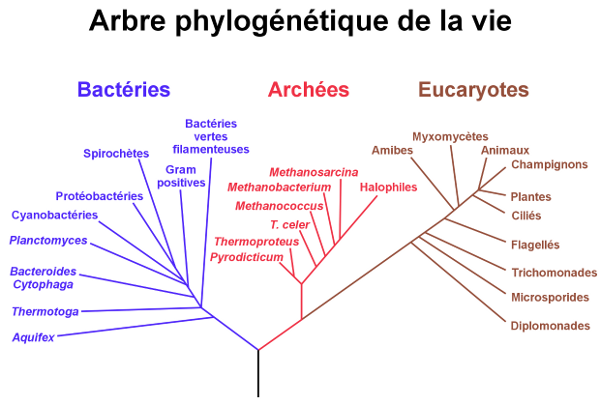
L’arbre du vivant se compose de trois domaines » : les êtres vivants dont l’ADN est enfermé dans un noyau, ou « eucaryotes », les bactéries, et les archées. Les archées sont des unicellulaires qui ressemblent à des bactéries. C’est Carl Woese qui les a, le premier, définies comme un groupe à part4. La plupart d’entre elles vivent dans des milieux extrêmes : chauds, acides, ou salés5. Il y en a dans les sources d’eau chaude, ou dans... notre système digestif6. Ceci nous amène à notre première observation : il n’y a pas de « direction » de l’évolution vers la pluricellularité, contrairement à l’idée reçue qui voudrait que l’évolution se fasse du « simple » au « complexe », avec en prime l’idée que le « complexe » correspond à « ce qui nous ressemble le plus, à nous autres les humains », pluricellulaire avec des cellules spécialisées en organes. D’une part, les neufs dixièmes de l’arbre sont des unicellulaires et d’autre part, ils sont loin d’être « simples » ! Même si aux yeux du profane, une bactérie ne ressemble à rien de plus qu’une autre bactérie, leur biochimie regorge d’inventions ingénieuses, diverses, originales et complexes, et n’a finalement rien à envier à celle des animaux ou des plantes.
Reprenons notre arbre. Il faut noter que, même si cela ne se voit pas au premier abord, l’arbre du vivant en général est un peu compliqué par quelques particularités liées aux bactéries. En effet, certaines de celles-ci se sont liées fortement avec les eucaryotes, en vivant en symbiose avec eux, à l’intérieur des cellules. Ces bactéries ont donné des organites aux rôles essentiels : mitochondries pour la respiration, chloroplastes pour la photosynthèse des végétaux... Il faut donc considérer l’évolution de ces organites séparément de celle de leurs hôtes eucaryotes. En outre, les bactéries ont l’habitude de s’échanger du matériel génétique entre individus (l’art de pratiquer le sexe en dehors de la reproduction). Ces « transferts horizontaux » (voir encadré 1.12 ) font parfois ressembler leurs branches de l’arbre à un réseau davantage qu’à un buisson.
Nous allons maintenant regarder plus attentivement la branche des eucaryotes. Les eucaryotes, ou « organismes dont les cellules comportent un noyau », se sont, comme les bactéries et les archées de leur côté, largement diversifiés. Dans les grands groupes ainsi apparus, on peut citer les animaux, les champignons, les végétaux, mais il en existe beaucoup d’autres, incluant notamment des organismes unicellulaires. Et il faut savoir que les champignons sont classés plus près des animaux que des végétaux, justement suite aux analyses génétiques.
La science qui consiste à explorer les liens exacts entre toutes les branches de l’arbre du vivant, une sorte de généalogie du vivant, s’appelle la phylogénie. Les découvertes récentes dans cette science nouvelle qu’est la phylogénie ont beaucoup bouleversé la conception traditionnelle du vivant, en seulement une trentaine d’années, même si certains des anciens grands biologistes avaient remarquablement bien déduit certains liens de parenté entre les organismes.
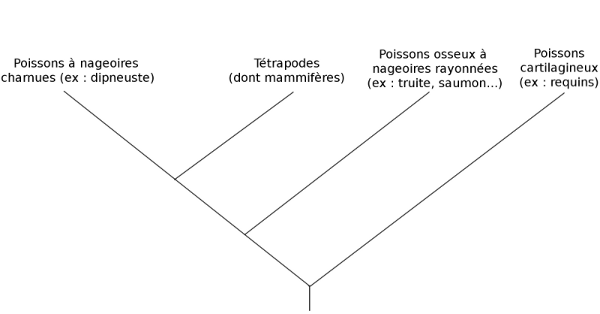
Nous savons maintenant que des termes comme « reptile », « poisson » ou « algue », n’ont pas de sens en phylogénie. En effet, si l’on prend les reptiles, il faut y inclure les dinosaures... Mais comme les analyses génétiques et paléontologiques ont également montré que les oiseaux étaient les descendants actuels d’une famille de dinosaures, cela signifie qu’il faut en toute logique inclure les oiseaux dans les reptiles ! Un nouveau groupe a donc été créé, celui des « archosauriens », pour désigner l’ensemble « dinosaures+oiseaux+crocodiles ».
Pour les « poissons », le mieux est de regarder l’arbre suivant pour comprendre le problème :
Et l’homme» ? L’homme est un animal, il vient donc se placer dans la branche des eucaryotes, au niveau des mammifères, juste à côté des chimpanzés et bonobos, dans la branche des primates.
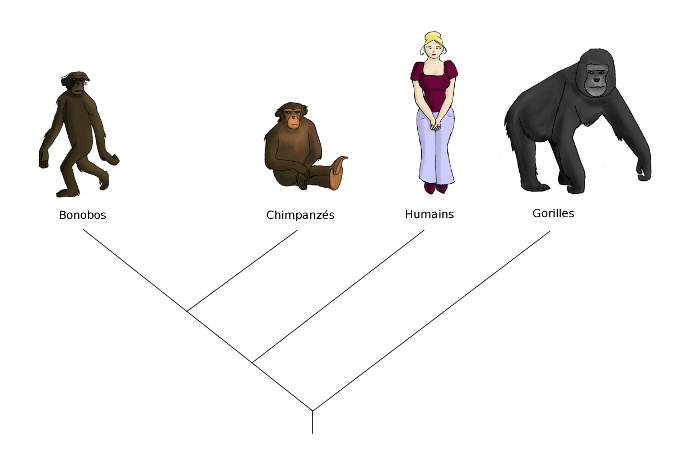
Nous avons commencé ce chapitre en parlant des races de chiens. Peut-on parler de races d’hommes ? Tout le monde peut voir que nous sommes très différents les uns des autres. Il y a des noirs, des blancs... En fait, tout serait simple s’il n’y avait que des noirs et des blancs, mais il y a des café au lait, des bronzés, des plus ou moins jaunes, des rougeâtres, bref, tout un continuum d’intermédiaires que personne ne sera jamais d’accord pour classer « noir » ou « blanc ». Rien que sur la couleur de la peau, on pourrait faire quatre, cinq, cent, mille « races », et encore, il y en aurait toujours qu’on ne saurait pas classer.
Et il n’y a pas que la couleur de la peau, il y a la forme du nez, des yeux, des cheveux... et surtout tout un tas de caractères invisibles, comme les groupes sanguins. Alors en admettant que l’on arrive à définir un certain nombre de groupes, en se basant sur la couleur de la peau, que se passera-t-il si on en fait d’autres en se basant sur les groupes sanguins ? Il seront différents, et ne se superposeront pas. On peut toujours choisir un ou deux (ou dix, ou mille) critères arbitraires, pour essayer de classifier les populations humaines, mais si l’on en change, on se rend compte que, peu ou prou, la classification a changé aussi.
Donc, que choisir ? Qu’est ce qui est le plus valable ? Les groupes sanguins, ou la couleur de la peau ? Pour les chiens, c’est facile, ce sont les éleveurs qui décident ce qui est important, en fonction de leurs envies. Mais ça ne veut pas dire qu’un caractère ait biologiquement plus d’importance qu’un autre. En plus, ces critères ne reflètent pas toujours les véritables filiations. Par exemple, plusieurs populations noires de peau pourraient être plus différentes entre elles (si on utilise une mesure sur d’autres gènes que ceux de la couleur de la peau) qu’une très foncée et une très claire. La coloration foncée de la peau est due à la sélection par l’exposition au soleil, qui fait que toutes les personnes habitant dans des régions ensoleillées vont avoir tendance à mourir tôt de problèmes de peau si elles n’ont pas assez de protection mélanique (le pigment noir de la peau, qui protège des UV solaires). Ainsi, deux populations foncées pourraient habiter côte à côte parce qu’elles sont issues toutes les deux de la même population-mère foncée, ou bien l’une est immigrante, mais que seuls ses membres les plus foncés aient été sélectionnés (c’est à dire qu’ils aient eu plus d’enfants que les autres, mourant moins jeunes).
Plus fort encore : la variation à l’intérieur d’un groupe est toujours plus élevée que la variation entre les groupes. C’est à dire que pour un gène donné, vous n’avez généralement pas davantage de chances de trouver une personne ayant les mêmes allèles que vous à l’intérieur de votre « groupe ethnique » qu’à l’extérieur de celui-ci7. L’illustration flagrante de cet état de faits est la recherche de donneurs dans le cas d’une greffe d’organe : si l’on ne trouve personne de compatible dans la famille proche du receveur, alors peut chercher directement dans l’ensemble de la population humaine, car les probabilités de trouver un donneur deviennent identiques quelle que soit la région du monde où l’on cherche.
Ainsi, pour définir de façon absolue des populations humaines que personne ne puisse contester, il faudrait compiler tous les caractères possibles. 30 000 gènes, certains très polymorphes, plus les séquences d’ADN non-codantes qui servent à mesurer le polymorphisme... On se retrouve alors avec un seul individu par race ! Nous sommes tous parents, mais tous différents.
Pourtant, l’homme est une espèce qui, comparée à d’autres (notamment au chimpanzé), n’a pas beaucoup de diversité génétique...
Il est donc impossible de biologiquement catégoriser des « races » humaines. Cela ne signifie pas que nous soyons tous identiques, au contraire, mais qu’il n’y a pas de limite nette quand on définit des populations. Alors, on peut très bien le faire pour les besoins d’une étude, mais en réalité, chaque étude ne fait qu’user une classification arbitraire qui arrange ses concepteurs, et elles sont toutes potentiellement différentes. Comme chacun l’accepte, cela ne pose pas de problème. Les médecins s’en servent d’ailleurs parfois pour faciliter certains diagnostics par exemple (certaines maladies génétiques sont plus présentes dans certaines régions du monde, ce qui ne signifie pas qu’on ne puisse les trouver ailleurs), ou affiner le dosage de certains médicaments afin de l’adapter à chacun selon son ascendance, en gardant à l’esprit les sévères limites de cette méthode8.
Au final, ce que nous apprend la génétique des populations sur l’humain, c’est que nous sommes beaucoup plus proches parents les uns des autres que ce que l’on croit habituellement9. Dès que l’on remonte de quelques générations dans le passé, nos arbres généalogiques ne ressemblent plus vraiment à des buissons, mais davantage à des filets, car nous partageons finalement tous le même nombre limité d’ancêtres.
Revenons à l’arbre du vivant. Il a donc une échelle de temps : on part de l’ancêtre, et on arrive aux espèces actuelles, qui sont toutes placées à égalité sur la ligne du présent.
Vous aurez compris que contrairement à ce que beaucoup d’entre nous ont appris à l’école, le vivant ne se représente pas sous forme d’une échelle, avec les « primitifs » en bas et les « évolués » en haut (et l’homme au sommet), car l’idée de « progrès » ne fait pas partie du concept d’évolution biologique.
On doit examiner les caractères un par un. Il y a les « ancestraux », qui étaient déjà portés par les ancêtres, dans le passé, et les « dérivés », qui ne l’étaient pas, et sont nouveaux. Un même organisme peut donc porter des caractères ancestraux et des caractères dérivés. Par exemple nous portons le caractère ancestral de la main à cinq doigts (on peut opposer le cheval, qui a un caractère dérivé d’une main à un seul doigt central, mais dont les ancêtres avaient aussi cinq doigts), et le caractère dérivé du pouce opposable. Quoi qu’il en soit, évitons d’employer le terme « primitif », que ce soit pour parler d’un caractère ou d’un organisme entier, qui est péjoratif : ce n’est pas parce qu’un système est vieux qu’il est inefficace (au contraire, si il est encore là de nos jours, c’est bien qu’il est efficace), ou qu’il ne l’a pas été dans un contexte environnemental donné, même si celui-ci est passé et n’existe plus. Chaque organisme est ou a été adapté à son milieu à un temps donné.
De la même manière, la « complexité » n’est pas un gage d’efficacité. De nombreux organismes sont restés simples au cours de leur évolution, d’autres se sont complexifiés. D’autres encore ont même connu des phases de complexification/simplification, comme les parasites qui peuvent se simplifier en perdant des fonction biologiques, effectuées par leur hôte à leur place. Chacun est adapté à son environnement propre.
En conclusion, nous pouvons dire que vu sous cet angle, il n’existe pas d’organisme plus « évolué » qu’un autre, car tout le monde a disposé du même temps d’évolution, l’homme, comme le poisson rouge, le brin d’herbe ou les bactéries.
3.5 L’évolution dans le temps
Nous avons pu voir quels étaient les quatre moteurs de l’évolution. Mais combien de temps cela prend-t-il, la spéciation, la différentiation des espèces ? C’est variable, car plusieurs facteurs sont en cause, tels que le temps de génération ou la nature des mutations, mais c’est en moyenne de l’ordre du million d’année, sachant que c’est parfois bien plus rapide... ou plus lent.
Chez les chiens, plusieurs milliers d’années de sélection n’ont pas permis de provoquer de spéciation, quoique l’on puisse se poser la question de la possibilité de croisement entre un mastiff et un chihuahua... C’est parce que cette sélection est un processus lent, qui ne touche que quelques caractères à la fois. Pour prendre un exemple chez une autre espèce, le chat, la photo ci-dessous vous montre l’évolution morphologique du profil du visage chez le siamois, en vingt ans de sélection.

Il est par conséquent impossible d’observer de son vivant de grosses modifications évolutives chez une espèce. Par contre, on peut voir se mettre en place des débuts de spéciation (sous forme par exemple d’isolement reproducteur partiel) chez des espèces qui ont des temps de génération rapides.
résumé!
Nous avons donc vu que l’évolution des espèces est explicable grâce à quatre phénomènes :
| – | les mutations, qui créent de la diversité génétique |
| – | la dérive génétique, qui réduit aléatoirement cette diversité |
| – | la sélection naturelle, qui réduit aussi la diversité, mais cette fois en fonction de l’environnement |
| – | la migration : les flux de gènes entre populations permettent d’augmenter une diversité perdue par dérive ou sélection. |
Il n’y a pas
besoin d’autre chose pour expliquer l’évolution du vivant dans le sens
où ces mécanismes conjugués sont capables d’entraîner les phénomènes
macro-évolutifs comme la spéciation ou les extinctions, pour peu qu’on
leur laisse suffisamment de temps.
Grâce à l’étude
de ces mécanismes, on peut construire des arbres pour représenter les
degrés d’apparentement entre espèces. En connaissant le rythme des
mutations, on peut même avoir une idée de l’âge de l’ancêtre de deux espèces.
1. Mayr E. 1963. Animal Species and Evolution. Cambridge (MA) : Harvard University Press.
2. Pour avoir une explication détaillée de l’arbre et de ses évolutions, je vous suggère de lire la « Classification phylogénétique du vivant » de Guillaume Lecointre et Hervé Le Guyader, aux éditions Belin
3. Source : Wikipedia, http://commons.wikimedia.org/wiki/Image:Phylogenetictree.svg, licence : Domaine Public.
4. Woese CR, Fox GE. Phylogenetic structure of the prokaryotic domain : the primary kingdoms. Proc Natl Acad Sci U S A. 1977 Nov ; 74(11) : 5088-90.
5. Pour en savoir davantage sur la découverte et le mode de vie de ces très passionnants organismes, je vous conseille la lecture de l’ouvrage suivant : « Les microbes de l’enfer », Patrick Forterre, ed Belin/Pour la Science
6. Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE, Relman DA. Diversity of the human intestinal microbial flora. Science. 2005 Jun 10 ; 308(5728) : 1635-8.
7. Rosenberg
NA, Pritchard JK, Weber JL, Cann HM, Kidd KK, Zhivotovsky LA, Feldman
MW : Genetic structure of human populations. Science. 2002, 298 : 2381-2385.
8. Lee SS, Mountain J, Koenig B, Altman R, Brown M, Camarillo A, Cavalli-Sforza L, Cho M, Eberhardt J, Feldman M, Ford R, Greely H, King R, Markus H, Satz D, Snipp M, Steele C, Underhill P. The ethics of characterizing difference : guiding principles on using racial categories in human genetics. Genome Biol. 2008 ; 9(7) : 404.
9. Rohde DL, Olson S, Chang JT. Modelling the recent common ancestry of all living humans. Nature. 2004 Sep 30 ; 431(7008) : 562-6.