L'évolution du vivant expliquée à ma boulangère
Chapitre 1
1.1 Un exemple : le chien
Pour comprendre les mécanismes de l’évolution, rien n’est plus simple que de regarder les animaux domestiques. Darwin avait d’ailleurs étudié, entre autres, les pigeons domestiques.
Le chien est aussi un bon exemple. L’archéologie permet de trouver des traces de domestication il y a au moins 17 000 ans1. Lorsque les premiers chiens furent domestiqués, ils ressemblaient vraisemblablement beaucoup au loup, dont ils sont issus, ainsi que nous l’indiquent les dernières recherches effectuées à ce sujet (bien qu’il existe également des hypothèses suggérant que l’origine du chien soit aussi liée à une domestication du coyote et/ou du chacal). Si l’on regarde l’ensemble des races actuelles, par contre, il faut parfois beaucoup d’imagination pour y voir encore un semblant de loup.
 |
Pensez au berger allemand... là, ça va encore. Mais le Saint-Bernard, le lévrier, le chihuahua, le basset et le caniche nain ? Comment toutes ces races sont-elles apparues ?

Reprenons notre premier chien domestique, ou plutôt, notre première population de chiens domestiques. Imaginez les premiers éleveurs, avec leurs premiers élevages, en famille. Les premiers élevages de chiens ont dû être relativement consanguins (la consanguinité s’obtient par croisement d’individus apparentés, comme les frères et les sœurs, les cousins etc). Lorsque de nouveaux caractères ont été révélés chez certains chiots (nouvelles couleurs, modification de la longueur des poils etc...), les premiers éleveurs ont réussi à rendre leurs caractères préférés plus fréquents, en croisant entre eux des chiens choisis pour ces particularités. Avec le temps, ils ont réussi à les fixer, c’est à dire à avoir des chiens portant chacun les caractères désirés.

Regardons maintenant ce qui s’est passé au niveau génétique. Vous
n’êtes pas tout à fait sûr de savoir ce qu’est, concrètement, un gène ?
Allez donc jeter un œil à l’encadré ci-dessous...
Concepts basiques
Les gènes
Les êtres
vivants sont constitués de cellules. Ils peuvent en compter plusieurs,
voire plusieurs dizaines, centaines, milliers, ou milliards - comme nous
autres les animaux - ou une seule, comme les levures, ou les bactéries.
Chaque cellule
vivante est différente des autres, et a des particularités métaboliques
ou physiologiques. Comment une cellule « sait »-elle comment elle doit
être, et comment elle doit se comporter ? Pourquoi une bactérie ne peut-elle pas se transformer
subitement en autre chose ?
Dans chaque
cellule, il y a son « mode d’emploi », ou son « plan ». C’est de
l’information que chaque cellule porte en elle et qui détermine son
aspect, son fonctionnement, ses réactions à son environnement.
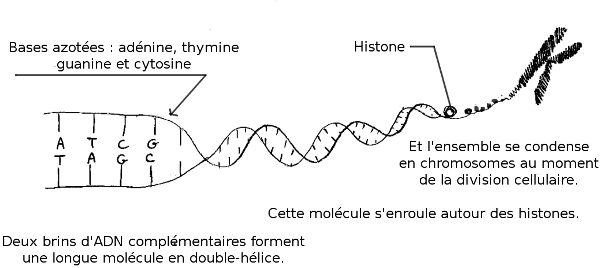
Cette
information est portée par de longues molécules d’ADN. Une molécule
d’ADN forme ce que l’on appelle un chromosome. Chez les espèces du groupe des eucaryotes (animaux, végétaux, et champignons entre autres), les chromosomes sont enfermés dans un
compartiment spécial de la cellule, que l’on nomme « noyau ». Chez les bactéries et les archées* (nous reviendrons plus tard sur les archées), les chromosomes ne sont pas séparés du reste.
L’information
portée par la molécule d’ADN est codée dans un alphabet à 4 lettres,
les bases azotées : thymine, guanine, adénine et cytosine. La suite de
ces « bases » forme une sorte de « phrase » que l’on nomme « séquence ». Les
gènes sont des informations portées par des fragments d’ADN. Ils sont
le plan de fabrication de protéines. On dit qu’ils « codent » pour des
protéines. La cellule les fabrique en « lisant » l’ADN. Chaque chromosome peut contenir des centaines de gènes.
On peut donc dire que l’information génétique est lue par le système physique que constitue la
cellule, l’individu. L’information contenue dans le « système de
lecture » lui-même est appelée « information épigénétique » (voir encadré suivant).
Et entre les
gènes ? Il y a des séquences de régulation des gènes, mais aussi des
séquences qui ne semblent pas avoir d’utilité particulière, comme des
reliques de virus, des séquences répétées etc. Les tests de paternité sont basés sur
l’analyse de telles séquences répétées que l’on nomme
« microsatellites », très variables d’un individu à l’autre.
Dans les
organismes pluricellulaires, chaque cellule porte exactement la même
information génétique (à quelques mutations près, éventuellement, ce
qui la plupart du temps n’a pas de grande conséquence, sauf dans des cas particuliers, par exemple quand elles touchent des gènes susceptibles de causer des cancers). Pourquoi, dès lors, toutes nos
cellules ne sont-elles pas rigoureusement identiques les unes aux autres ? Si
nous avons des cellules sanguines, des neurones, des cellules
musculaires etc., c’est parce que l’expression de l’information
génétique peut être régulée par des facteurs extérieurs. C’est une part
de ce que l’on appelle la « plasticité phénotypique », la capacité d’un génome à produire un « phénotype » (ensemble des traits observables de la cellule, l’organisme) différent en fonction de l’environnement. La cellule-œuf
initiale, qui résulte de la fusion de l’ovule et du spermatozoïde,
contient en elle toutes les informations pour faire toutes les cellules
du corps humain. À partir de cellules embryonnaires « totipotentes », c’est à dire « ayant toutes les possibilités », tous les types de
cellules pourront se différencier, en fonction des signaux reçus des
autres cellules, de l’environnement, qui vont moduler l’expression des
gènes.
La science qui étudie ces mécanismes de différentiation des cellules se nomme biologie du développement.
Chaque chien possède des gènes particuliers, qui vont déterminer la couleur de son poil, sa longueur, mais aussi la forme de son corps, de sa mâchoire, sa taille ou la longueur de ses pattes (et bien d’autres choses encore). On dit qu’un gène « code » pour un caractère donné. Généralement, un même caractère peut être sous l’influence de plusieurs gènes, et il arrive aussi qu’un gène joue un rôle dans plusieurs caractères, par exemple en provoquant la fabrication d’une molécule qui sera utilisée dans plusieurs organes. Bien sûr certains caractères ne sont pas déterminés uniquement par les gènes, les facteurs environnementaux ont aussi leur mot à dire (on retrouve l’idée de « plasticité phénotypique » vue dans l’encadré sur les gènes). Par exemple, un chien mal nourri ne sera jamais aussi grand que s’il était bien nourri. Mais c’est une autre histoire, car de telles caractéristiques dues à l’environnement ne seront pas transmises à la descendance (mis à part dans les cas particuliers des facteurs épigénétiques - voir encadré 1.1 ) : un chien de petite taille car mal nourri pendant sa croissance pourra avoir des chiots de grande taille s’ils disposent d’une alimentation correcte.
Pour aller plus loin
Les effets épigénétiques et les effets maternels
Il
serait trop simple que l’information héritable d’une génération à
l’autre ne soit portée que par la molécule d’ADN, à l’exception de tout
autre moyen. On sait maintenant que des molécules produites au cours de
la vie de la cellule peuvent influencer l’expression des gènes des cellules-filles. De l’information est portée dans le « système de lecture de l’ADN » que constitue la cellule.
C’est
ce qui explique notamment pourquoi d’une cellule-œuf, on peut arriver à
obtenir tout un tas de cellules, qui, bien que portant la même
information génétique, sont toutes différentes (cellule musculaire,
cellule nerveuse, etc...). Cela explique également comment ces cellules peuvent se diviser et donner des cellules-filles conservant leur spécialisation.
Ainsi,
même si un gène est présent et fonctionnel dans une cellule, son
expression peut être modulée par des molécules acquises au cours de la
vie de l’individu. Certains gènes sont activés, d’autres inhibés, etc.
Bien entendu, ces phénomènes épigénétiques sont réversibles (au moins
en théorie, même si dans la pratique les cellules différenciées ne
reviennent parfois plus en arrière, au moins chez les animaux), puisqu’ils ne
touchent pas au génome en lui-même.
Examinons
une des possibilités : imaginez que certaines de ces molécules soient
présentes dans l’ovule, et qu’elles puissent changer en fonction du
mode de vie de la femelle qui a produit cet ovule... C’est alors
l’histoire de vie de la mère qui pourra influer l’expression des gènes
de son descendant. Ce sont les effets maternels. Certains croisements peuvent donc avoir des résultats inattendus, à cause de ce type de mécanisme.
Attention
toutefois, n’oublions pas que ces phénomènes sont réversibles... c’est
à dire que ce ne sont pas les gènes en eux-mêmes qui sont modifiés, et
que d’une génération à l’autre la situation peut potentiellement s’inverser, éventuellement en une seule génération. Par
conséquent, ces phénomènes ne sont pas à rapprocher de l’idée de
l’hérédité des caractères acquis, notamment avancée par Lamarck en son
temps. Cette idée suggère que les modifications morphologiques acquises par l’individu pendant sa vie, telle par exemple une forte musculature suite à un entraînement prolongé, se transmettraient à sa descendance. Cette idée reste fausse. Une personne fortement musclée pour cause d’une pratique d’activité physique intense n’a pas plus de chances que les autres d’avoir des enfants musclés si ceux-ci ne font pas particulièrement de sport.
Dans notre population « ancestrale » de chiens, tous avaient à peu près
le même aspect (en tous cas, il y avait beaucoup moins de différences
entre eux qu’entre le Saint-Bernard et le chihuahua, tout le monde
ressemblait au loup). Mais avaient-ils les mêmes gènes ? Non. Ce n’est
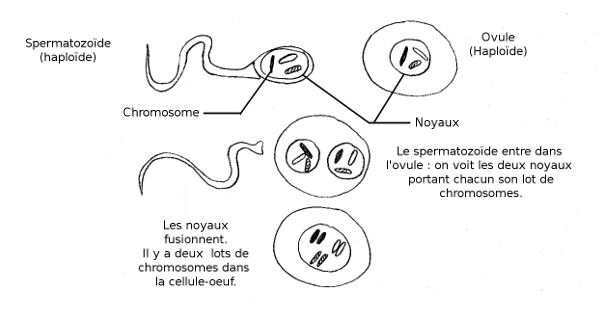
pas obligatoire, loin de là. Les chiens, comme tous les mammifères,
sont diploïdes. C’est à dire que tous leurs
chromosomes vont par paire - un chromosome est un long filament d’ADN, enroulé sur
lui-même et empaqueté dans des protéines quand la cellule se divise, déroulé quand elle l’utilise pour fabriquer d’autres protéines. Pour chaque paire, un des chromosomes vient de la mère (via l’ovule), et un autre vient du père
(via le spermatozoïde). Ces deux chromosomes sont dits« homologues » car ils contiennent la même suite de gènes.
Les chiens, comme nous et beaucoup d’autres organismes, ont ainsi chaque gène en double exemplaire.
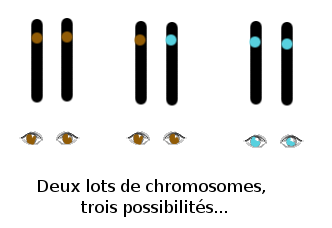
Vous pouvez donc imaginer la situation : l’individu peut porter deux fois le même exemplaire de chaque gène, ou bien alors deux exemplaires différents (on parle d’« allèles »). Dans le premier cas, on dit que l’individu est « homozygote » pour le gène considéré, dans deuxième cas, on dit qu’il est « hétérozygote ».
Ainsi, chez les humains, un bébé ayant un parent à peau claire et un parent à peau sombre héritera, pour chaque gène impliqué dans la couleur de la peau (attention, il y en a plusieurs !), d’un allèle de son père et d’un autre de sa mère, et aura le plus souvent une peau de couleur intermédiaire.
Un phénotype (trait ou ensemble de traits, observables, physiques, d’un organisme) peut aussi être dominant sur l’autre (qui est alors dit « récessif ») c’est à dire qu’il en masque l’expression. Pour la couleur des yeux, par exemple : si le phénotype « couleur marron » est dominant sur le phénotype « couleur bleue », alors l’individu qui possède un allèle de chaque aura les yeux... marron. C’est bien, je vois que vous suivez.
Mais attention, la couleur des yeux n’est en réalité pas soumise à un seul, mais à plusieurs gènes, chacun possédant potentiellement plusieurs allèles... Donc, si vous désirez en savoir un peu plus sur la génétique, et la prévision des résultats des croisements, toujours sur l’exemple de la couleur des yeux, l’encadré 1.1 est là pour vous.
Concepts basiques
Peut-on prévoir les résultats d’un croisement ?
Tout à fait, à
condition bien sûr que le caractère que l’on étudie soit génétiquement
transmissible, et que ce déterminisme soit connu. C’est d’ailleurs une des bases de la génétique, initiée
par le moine Mendel. Chaque parent transmet, via ses gamètes (ovule ou spermatozoïde chez les animaux), un de ses
allèles à son descendant. Chaque allèle a donc une chance sur deux
d’être transmis.
Si l’on sait
quels allèles possèdent les parents, on peut prévoir ceux des
descendants, en terme de probabilités. Inversement, voir les allèles
d’une descendance permet de déduire le génotype des parents (quels
allèles ils possèdent).
Reprenons un
exemple : celui de deux couleurs d’yeux, bleu et marron. Ces couleurs
sont dues à la mélanine, pigment marron foncé, qui, dans un cas, sera
présente dans les couches profondes de l’iris seulement (œil bleu), et
dans l’autre, aussi en surface (œil marron).2
Chez un
individu aux yeux bleus, la mélanine n’est pas produite/amenée dans les
couches superficielles de l’iris, car une des enzymes, de synthèse
ou de transport du pigment, ne fonctionne pas, ou est
absente. Cependant, généralement, si un individu possède un allèle
efficace de cette enzyme, et aussi un allèle défectueux (hétérozygote,
donc), il pourra quand même avoir des yeux marrons, puisque un allèle
suffit à fabriquer l’enzyme. Par exemple, l’albinisme étant un défaut de fabrication d’une enzyme servant à synthétiser la mélanine, si une personne est hétérozygote, on la qualifiera de «porteuse », car elle aura de la mélanine mais ses enfants pourront être albinos si l’autre parent est hétérozygote pour le même gène également.
 |
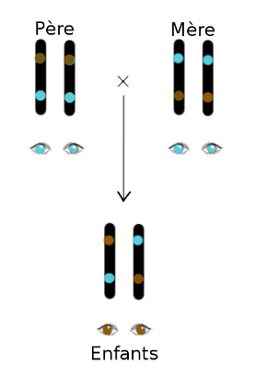
| Figure 1.6 Exemple simple d’un parent aux yeux bleus, et de l’autre aux yeux marrons, hétérozygote (portant un allèle pour le bleu, et l’autre pour le marron). |
L’exemple de la figure 1.6 représente un cas classique. Pour être sûrs que le principe soit bien compris, prenons maintenant l’exemple d’un autre couple. Imaginons deux parents aux yeux marrons, hétérozygotes (un allèle
fonctionnel, et pas l’autre). Ils auront chacun une chance sur deux de
transmettre à leurs enfants chacun de leurs allèles (voir fig 1.7 ). Ils auront donc
une chance sur quatre d’avoir un enfant homozygote aux yeux marrons,
deux chances sur quatre d’avoir un enfant hétérozygote aux yeux
marrons, et une chance sur quatre que l’enfant soit homozygote aux yeux
bleus. Vous suivez toujours ?
 |
| Figure 1.7 Tableau de croisement : la couleur des yeux, version très simple avec un gène à deux allèles, parents hétéroygotes. |
Les deux exemples précédents sont simples, mais la réalité est plus compliquée, car ce n’est pas un seul gène, mais plusieurs qui sont impliqués dans la couleur des yeux3. Pour nous donner une idée des mécanismes, intéressons-nous
à la synthèse du pigment. La mélanine est fabriquée en plusieurs
étapes. De manière générale, une chaîne de synthèse se déroule comme suit : une première molécule est synthétisée (ou absorbée par la nourriture), puis intervient dessus une autre molécule nommée enzyme, qui transformera par réaction chimique notre première molécule en une deuxième,
que l’on l’appellera, après réaction, « produit ». Les enzymes, souvent des protéines, sont spécialisées dans l’accélération des réactions chimiques, on les appelle des « catalyseurs ». Plusieurs autres enzymes interviennent ensuite successivement sur le produit ainsi formé, le transformant, dans notre cas, en mélanine, produit final.
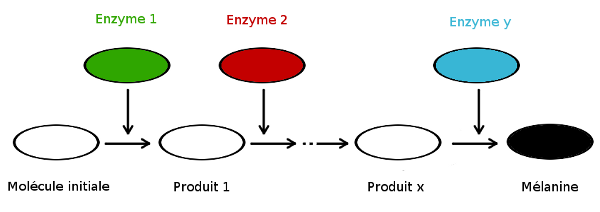
Les enzymes sont codées chacune par un gène précis.
Mais la mélanine
n’est pas forcément produite directement là où elle est exposée : elle
peut aussi avoir besoin de migrer, ce qui est permis par d’autres
molécules, sur lesquelles interviennent d’autres enzymes. Ça devient
compliqué, parce que des tas d’enzymes différentes fonctionnent ensemble, donc il y a autant de chances que la chaîne soit interrompue quelque part !
C’est ce type de mécanisme qui explique qu’il arrive aussi que des
parents aux yeux bleus aient des enfants aux yeux marron. Avant de
conclure un échange de bébé à la maternité, ou une intervention du
facteur, il convient de bien comprendre l’histoire de la mélanine.
Si le père ne peut pas
fabriquer la première enzyme, et que la mère ne peut pas fabriquer la
troisième, l’enfant sera hétérozygote pour chacun des gènes codant pour
ces enzymes... et donc, il pourra, lui, les produire !
Et ses yeux seront... marrons. Cela se nomme la « complémentation ».
La génétique de la couleur des yeux est donc très compliquée. Récapitulons.
D’abord, il n’y
a pas que deux couleurs, mais des dizaines : bleu, gris, vert (et le vert peut être dû à un autre pigment que la mélanine, nommé lipofuscine), noir, brun,
noisette, avec toutes les nuances possibles. Ensuite, on connaît
quelques gènes qui sont à coup sûr impliqués dans certaines de ces
couleurs, mais ils n’expliquent qu’une part de la variabilité à
l’intérieur de la population humaine. À l’heure actuelle, si les mécanismes généraux sont connus, il manque encore bien des détails.
En outre, la
synthèse des pigments n’est pas seule en cause : ils peuvent
être synthétisés, mais pas « déposés » dans les différentes structures de
l’iris, ce qui influe encore sur la couleur. Et la densité de cellules entre les couches profondes et superficielles de l’iris peut également influer cette couleur.
Enfin, si l’on
prend en compte que plusieurs mécanismes (dominance, codominance,
complémentation) peuvent jouer au niveau des liens entre les allèles et les gènes, on comprend qu’il n’est pas rare de se retrouver dans un cas qui ne colle pas avec les exemples
habituels (naissance d’enfants avec une couleur des yeux inattendue).
Moralité : oui,
on peut souvent prévoir les résultats des croisements, mais il faut
prendre toutes les précautions et bien penser aux différents mécanismes
impliqués. On aura donc un résultat sous forme de probabilités : x% de
chances d’obtenir tel phénotype.
Les éleveurs et
les généticiens ont l’habitude de jongler avec de telles probabilités.
Cependant, le travail est parfois très ardu.
Revenons à nos chiens. Si nous avons des chiens portant des allèles codant pour des phénotypes récessifs rares, cela ne se voit pas, parce que ces allèles se trouvent toujours associés à d’autres, codant pour un phénotype dominant. Par contre, quand, par hasard, sont croisés deux individus portant le même allèle responsable de ce phénotype, il est possible que naisse un chiot homozygote pour cet allèle particulier (portant donc deux exemplaires identiques du gène considéré, comme la personne aux yeux bleus de notre exemple). Ce chiot exprime le phénotype rare, dont l’existence est ainsi révélée.
En croisant les homozygotes pour cet allèle, avec les hétérozygotes (souvent les parents ou les frères et sœurs, dont on pense qu’ils peuvent porter une copie de cet allèle récessif), on finit par obtenir de plus en plus d’individus homozygotes pour l’allèle récessif. Et le tour est joué. Quelques milliers d’années plus tard, nous avons notre caniche nain, alors que les gènes donnant au loup des poils frisés, une petite taille, une robe blanche, un front bombé avec des grands yeux, sont extrêmement rares dans la nature ! Vous voyez également en quoi la consanguinité (résultant de croisement entre apparentés), recherchée volontairement ou non par les éleveurs, peut accélérer le processus.
1.2 Quels sont les mécanismes en jeu dans l’exemple du chien ?
Nous sommes partis d’une population apparemment assez homogène (de loups), pour arriver à une multitude de petites populations très différentes les unes des autres (les caniches, les bergers allemands, les rottweilers, les show show...).
La population de base possédait un certain polymorphisme génétique (c’est à dire, plusieurs allèles pour chaque gène codant pour les caractères qui nous intéressent, comme la couleur ou la longueur des poils). En divisant la population initiale en petites sous-populations avec chacune ses caractères particuliers, ce polymorphisme s’est trouvé fortement réduit dans chacune d’elle.
Mais si l’on décidait de mélanger toutes les races de chiens, et que l’on comptait les allèles présents, qu’observerait-on ? Ces allèles sont globalement les mêmes que dans la population ancestrale, mais dans des proportions différentes (à quelques mutations près, voir chapitre 2). Ceux qui étaient rares dans la population ancestrale sont devenus fréquents dans certaines populations actuelles (et on peut ajouter que des allèles peuvent même avoir disparu dans quelques populations).
C’est la conséquence de la sélection effectuée par les éleveurs. C’est une sélection artificielle.
Nous pouvons noter que la sélection peut faire perdre des allèles à une population. Pour les réintroduire, les éleveurs font parfois des croisements entre races, ce qui leur permet de « réinjecter » les allèles perdus dans leur population.
Mais laissons là nos chien un moment, et penchons nous sur ce qui se passe dans la nature.
1.3 Et dans la nature ?
Dans la nature, trois phénomènes agissent de concert sur la diversité génétique :
| – | La sélection naturelle, |
| – | La dérive génétique, |
| – | Les flux de gènes. |
a. La sélection naturelle
L’environnement exerce ce que l’on apppelle une « pression de sélection ». Dans un environnement donné, certains caractères représentent un avantage en facilitant la vie des individus qui les portent, d’autres non. Si ces caractères sont associés à des
allèles*, et donc transmissibles à la descendance, les allèles désavantageux vont devenir moins fréquents, jusqu’à éventuellement disparaître de la population, alors que d’autres vont mieux se
transmettre d’une génération à l’autre, et devenir plus fréquents.
Revenons un instant sur la notion d’allèle, ou de caractère, « avantageux » : cette question est très importante. Un caractère n’est jamais avantageux dans l’absolu. Par exemple, est-il avantageux ou pas d’avoir des poils longs ? En fait, ça
dépend. Si vous vivez au froid, oui, si vous vivez au chaud... pas toujours.
 |
Voyons maintenant les facteurs ce qui se cachent derrière le terme « environnement ». Non seulement
les espèces subissent des pressions de sélection de la part de
facteurs comme le climat, mais aussi de la part des autres espèces avec
lesquelles elles interagissent (pathogènes, parasites, prédateurs, compétiteurs, proies...). De véritables « courses aux armements » peuvent s’enclencher. Par exemple, la proie possédant un caractère lui permettant d’échapper au prédateur, comme un camouflage, contribuera à la sélection des prédateurs les plus à même de la détecter malgré son camouflage. Le prédateur, de son côté, en éliminant les mauvais camouflages, sélectionnera les plus efficaces. Ainsi, si un prédateur se nourrit de plusieurs espèces-proies, et que l’une d’elles dispose d’un
nouveau caractère pour échapper au prédateur, la prédation sur
elle-même diminuera, mais sur les autres espèces elle augmentera potentiellement en
conséquence. Les proies, comme les prédateurs, sont en compétition les uns avec les autres. Plusieurs points de vue sont donc possibles : relations prédateurs/proies, compétition prédateur/prédateur ou proie/proie. On parle alors de sélection inter-spécifique. Tout ceci s’ajoute à la compétition qui a lieu entre individus de la même espèce (sélection intra-spécifique), pour échapper aux prédateurs, aux pathogènes, ou trouver les meilleures proies.
Pire, un allèle peut très bien être favorable à un individu par rapport aux autres de son espèce, mais uniquement en présence d’un autre allèle (d’un autre gène, ou du même gène), et défavorable dans le cas contraire. Par exemple, un allèle codant pour des crocs pointus ne sera favorable qu’en présence d’allèles permettant la digestion de la viande, et d’allèles favorisant un comportement de chasse. L’environnement d’un gène est donc à prendre dans sa globalité : il s’agit de l’environnement extérieur à l’individu, mais aussi des autres gènes portés par l’individu !
Pour comprendre si un allèle est avantageux, il faut donc l’examiner à la fois dans le contexte environnemental de l’espèce, par rapport aux autres allèles du même gène, portés par les autres individus et dans le contexte génétique de l’individu qui le porte. Il est par conséquent très difficile de savoir si un caractère est plus « favorable » que d’autres. D’une part, cette notion est relative et variable en fonction des facteurs environnementaux, et d’autre part, de nombreux caractères sont, même dans un environnement donné, à la fois favorables et défavorables. Un exemple connu chez l’homme est celui de la mutation du facteur de coagulation V dite « Leiden », du nom de l’université où elle a été découverte4. Elle provoque une augmentation de la vitesse de coagulation du sang. Il a été montré que cette mutation pouvait augmenter la résistance à certaines infections5, mais d’un autre côté, elle provoque un risque accru de phlébites. Ce genre de caractère est dit soumis à un compromis. Il n’y a pas de « solution parfaite ». Pour mieux comprendre encore, un autre exemple, chez la mouche du vinaigre : il a été montré que la bonne mémoire avait un coût en terme de durée de vie6. Plus les mouches ont une bonne mémoire, ce qui est utile pour elles, pour trouver leur nourriture par exemple, moins elles vivent longtemps.
Et il faut aussi faire attention à un point important : en réalité, peu importe qu’un caractère soit défavorable à l’individu (ou « à l’espèce »). Un caractère est sélectionné simplement s’il est favorable à la reproduction de l’individu qui le porte (et s’il est héritable, bien entendu1.3 ). Il est facile de comprendre que si un caractère confère à un individu une vie fertile plus longue que celle de ses congénères, et donc plus de descendants sur l’ensemble de sa vie, il sera sélectionné si les conditions environnementales le permettent — pas trop de prédation sur les adultes par exemple. Mais inversement, si un allèle permet à un individu d’avoir une descendance fertile plus nombreuse que ses congénères dépourvus de l’allèle en question, et l’amène à mourir jeune, mais après sa reproduction, il sera sélectionné également ! Pourquoi ? Au court de sa courte vie, il aura eu davantage de descendants que les autres individus, même si la vie des autres a été plus longue.
Cet état de fait permet d’expliquer en partie le phénomène du vieillissement, dû à une accumulation, au cours des générations, d’allèles ayant un effet favorable ou neutre sur la reproduction, la bonne santé, dans la jeunesse, mais un effet plus ou moins délétère, quelque temps après l’âge de la première reproduction. Nous y reviendrons un peu plus tard.
Pourquoi de tels allèles sont-ils sélectionnés ? C’est mathématique : si un allèle permet d’avoir une descendance fertile plus nombreuse que les autres, étant transmis à cette même descendance, il lui permettra à son tour d’avoir une descendance plus nombreuse que les enfants de ceux qui possédaient un autre allèle. Comme aucune population ne peut croître indéfiniment, il y a une compétition entre les descendants de chaque génération, mais de part leur faculté à bien se reproduire, les individus portant l’allèle avantageux à la reproduction seront toujours plus nombreux, jusqu’à envahir la population. Cette notion est très importante. Vous pouvez faire une pause pour y réfléchir... Prenez un café, par exemple ;-). Vous l’avez bien mérité.
 |
La sélection naturelle est donc une force qui modifie, et souvent réduit la diversité génétique dans une population. Dès que les conditions environnementales changent, les pressions de sélection qui jouent sur la population se modifient. Cela entraîne des changements de fréquences dans les allèles, car les caractères qui étaient avantageux auparavant ne le sont plus forcément par la suite. Le nombre d’allèles a tendance à se réduire.
Attention toutefois. La sélection naturelle a de multiples facettes, et il est facile de faire des interprétations erronées. Par exemple, prenons le fameux exemple des girafes.
Dans une population ancestrale de girafes, à l’époque où leur cou était court (si vous vous demandez comment on sait que les ancêtres des girafes avaient le cou court, ce sera expliqué plus loin), il existait un certain polymorphisme des gènes qui codaient pour la longueur du cou. En gros, certaines avaient des cous un peu plus longs que d’autres. D’ailleurs, si vous regardez autour de vous, vous constaterez la même chose chez vos voisins : certains top-models ont des cous très longs, alors que d’autres personnes donnent l’impression de n’avoir pas de cou (et pourtant, tout le monde a le même nombre de vertèbres cervicales, même les girafes : sept. C’est le nombre de vertèbres cervicales des mammifères, qui est fixe, à l’exception des cas du lamantin et du paresseux78).
Revenons à nos girafes : les girafes qui naissaient avec un cou un peu plus long que les autres avaient une descendance plus nombreuse que celles à cou court, et de nos jours, toutes les girafes ont un long cou (ceci n’est valable que parce que l’effectif total des girafes ne peut pas croître, d’une génération à l’autre, jusqu’à l’infini). La question est : « Pourquoi le long cou a-t-il favorisé la reproduction des girafes ? »
| – | Peut-être parce qu’il leur permettait de brouter les feuilles plus haut dans les arbres, et qu’elles étaient donc mieux nourries, évitant la compétition avec les espèces à cou court mangeant la végétation basse (sélection naturelle stricte). |
| – | Peut-être parce que quand les girafes mâles se battent pour accéder aux femelles, elles utilisent leur cou... ce qui fait que les plus longs cous auraient gagné plus de combats, et donc obtenu la faveur de ces dames (sélection sexuelle – pour plus de détails sur cette forme de sélection, direction l’encadré sur la sélection sexuelle, ci-dessous !). |
| – | Peut-être que seul le hasard de la dérive génétique est en cause. |
 |
Il y a également des cas où la sélection va aider à maintenir la diversité, et pas trop la réduire. C’est le cas par exemple de la sélection dite « fréquence-dépendante ». Dans ce cas, un caractère est sélectionné quand il est rare. Cela peut par exemple expliquer pourquoi le système majeur d’histocompatibilité, le système qui permet l’identification du « soi » par le système immunitaire (et pose donc des soucis lors des greffes d’organes, car le système immunitaire du receveur reconnaît les nouvelles cellules comme étrangères), est composé de molécules si variées. En effet, les pathogènes (bactéries, virus etc...) auraient plus de difficulté à infecter un hôte au MHC rare, favorisant la survie et donc la reproduction de ce dernier9.
Pour aller plus loin
La sélection sexuelle
La sélection
sexuelle est un des aspects de la sélection naturelle. Avez-vous
remarqué que chez beaucoup d’espèces, notamment les oiseaux, les mâles
sont très différents des femelles ? Ils ont des couleurs vives, ou des
ornements divers. Des plumes colorées, des chants, des parades. Ils
présentent tout un tas de caractéristiques qui semblent en parfaite
contradiction avec ce qui serait convenable pour passer inaperçu des
prédateurs.
Chez certaines
espèces, ce sont les femelles qui portent ces jolis attributs voyants,
chez d’autres, ce sont les deux sexes, mais de manière générale, le
sexe qui exerce le « choix » le plus marqué, et donc qui est le moins
coloré/voyant/bruyant est celui qui investit le plus d’énergie dans la
production de chaque petit (par exemple, il va passer du temps à les
nourrir et à en prendre soin). Quand on investit beaucoup d’énergie dans un seul
petit, on a tendance à choisir plus sélectivement son partenaire que quand on investit dans un grand nombre de petits, mais peu d’énergie à chaque fois.
Et oui, les deux
parents n’ont pas forcément le même niveau d’investissement. Mais même
s’il l’est globalement, chacun aura quand même intérêt à ce que son
conjoint remplisse correctement sa part (de nourrissage des jeunes par
exemple), et même plus que correctement. Du coup, les intérêts des deux
sexes entrent facilement en conflit, voire divergent carrément,
entraînant des coévolutions, des courses entre stratégie et
contre-stratégie.
De ce point de
vue, chaque sexe peut ainsi être considéré comme une « ressource » pour
l’autre, en fonction de son investissement parental, de ses allèles, de
la disponibilité globale des partenaires. Comme ils sont plus gros, car pleins de ressources énergétiques pour le futur embryon, les ovules
produits par les femelles sont moins nombreux que les spermatozoïdes,
bien que des différences de sex-ratio (rapport entre le nombre de mâles et nombre de femelles dans une population) peuvent modifier la donne en
rendant les mâles rares. Chacun a donc intérêt à choisir la meilleure
« ressource », ou à être choisi par elle. Mais choisir un bon partenaire
est comme chacun sait un art difficile. Pourquoi le plus séduisant
est-il souvent le plus bizarre, ou le plus voyant ?
Tous ces
critères, de couleur, de chant, de parade, ne sont-ils pas apparemment
en contradiction totale avec la sélection naturelle ? Ces plumes,
cornes, bois, sont coûteux à produire, ils demandent de l’énergie,
énergie qui pourrait être dépensée à autre chose (le système
immunitaire par exemple). Ils sont coûteux à maintenir aussi, parce
qu’ils sont voyants et peuvent servir de point de repère aux
prédateurs, comme une grosse pancarte « Viande fraîche ici ! ».
Parfois, ces
ornements servent à la compétition intra-sexe, par exemple les combats
entre mâles. Pourtant, ils peuvent aussi ne s’adresser qu’aux
partenaires potentiels, pour les séduire, ou constituer à la fois un
avertissement aux concurrents et un signal de séduction, comme le chant
des oiseaux par exemple.
Il y a deux
grandes hypothèses pour expliquer l’évolution de tels caractères de
séduction : l’« emballement » de Fisher, et le principe du handicap de
Zahavi10.
Admettons que ce soit la femelle qui choisisse, ce qui
est le plus souvent le cas. D’après Fisher11, si une femelle préfère un mâle qui présente un certain
caractère, ses fils le posséderont également, si le caractère est héritable. Et si la préférence des femelles est aussi héritable, alors les filles préfèreront les mêmes types de mâles que leurs mères, sélectionnant au fur et à mesure des générations les mâles aux attributs les plus importants. Le phénomène s’entretient
de lui-même, s’emballant jusqu’à ce que le coût de production du
caractère devienne trop lourd.
Selon le
principe du handicap, c’est justement parce qu’ils sont coûteux dès le
début de processus (contrairement à l’idée de Fisher), que les
caractères de sélection sexuelle sont maintenus. Ils servent de point
de repère au sexe opposé, qui peut ainsi choisir le partenaire de meilleure qualité. À cause de ce coût, la tricherie n’est pas facile. Si un individu présente un caractère si coûteux à produire,
et qu’il est malgré tout là, à parader, c’est qu’il doit vraiment être
exceptionnel, non ?
Or, comment
choisir son partenaire, d’un point de vue évolutif ? Comment choisir
celui qui donnera la descendance la plus nombreuse et la plus
vigoureuse ? Selon la théorie des « bons gènes » (comprendre « les bons
allèles »), il faut être capable de trouver le partenaire qui a les
gènes les plus favorables en fonction de l’environnement dans lequel on se trouve. Le handicap
permet donc de « tester » le partenaire, et de trouver celui qui a de « bons
gènes ». Ainsi, vos petits seront de meilleure qualité, survivront mieux, et vos propres gènes seront le mieux disséminés, et donc
sélectionnés. Par conséquent, on peut dire que les femelles sont sélectionnées pour choisir les
« meilleurs mâles », et vice versa.
Bien sûr, la
triche est possible. Ainsi, un individu « moyen » pourrait investir toute
son énergie dans la production de signaux voyants, pour être choisi,
avoir une descendance nombreuse, avant de mourir épuisé. Ceci a été mis
en évidence chez ces petits poissons que sont les épinoches12. Le
partenaire « croit » choisir le meilleur, mais en fait il ne choisit
qu’un « moyen »...
Il y a encore
d’autres possibilités de sélection : certains allèles nécessitent, pour
être les plus favorables, les meilleurs, la présence d’allèles
« complémentaires ». C’est le cas de certains gènes du système
immunitaire. Les individus ne cherchent donc pas de « bons allèles »,
mais des allèles « complémentaires de ceux que j’ai ».
Dans tous les cas, disposer de signaux fiables permet d’identifier sûrement les partenaires potentiels porteurs de ces allèles.
C’est le rôle des signaux soumis à la sélection sexuelle : odeurs,
couleurs, chants, décorations diverses.
résumé!
Quels caractères sont susceptibles d’être sélectionnés ?
Tous ceux qui
sont héritables et qui jouent un rôle dans la capacité des individus à
se reproduire pour laisser une descendance féconde. Pour qu’une
sélection soit possible, il faut bien sûr que ces caractères soient
variables, c’est-à-dire que les gènes qui codent pour eux existent en
plusieurs allèles, certains laissant davantage de descendants que d’autres. Ces caractères peuvent être physiques, physiologiques, morphologiques, métaboliques, aussi bien que comportementaux.
b. La dérive génétique
La dérive génétique a le même type d’effet que la sélection naturelle. Elle modifie la fréquence des allèles dans la population, voire réduit la diversité allélique, pour la simple raison que certains individus ne contribuent pas, ou peu à la génération suivante, mais cette fois par pur hasard. En outre, dans le cas de la reproduction sexuée, les parents ne transmettent pas l’ensemble de leurs caractéristiques à leurs descendants, mais seulement quelques-unes (un allèle sur deux par descendant, nous y reviendrons plus bas). La principale différence entre la sélection et la dérive est donc que cette dernière s’effectue entièrement au hasard. Elle joue aussi un rôle dans l’évolution des espèces domestiques, comme le chien, notamment sur tous les caractères auxquels ne font pas attention les éleveurs, mais il est difficile de quantifier son rôle.
Quand on observe un phénomène, il peut appartenir à différentes catégories. Il peut se dérouler selon une loi ordonnée. On peut donc prévoir ce qui va se passer. Par exemple, un système ordonné est un jeu de cartes rangées dans l’ordre. Après chaque carte, on peut prévoir quelle sera la suivante.
Il y a aussi le système chaotique, dont une illustration est « l’effet papillon ». Dans le système du chaos déterministe, on pourrait prévoir ce qui se passera si on avait une connaissance infiniment précise de tous les paramètres initiaux, mais comme obtenir cette connaissance est impossible par définition (à cause de la notion d’« infini »), on se retrouve avec quelque chose d’en apparence imprédictible. On ne peut pas prévoir quel événement, parmi tous ceux qui sont possibles, va se produire.
Enfin, si on peut dégager une loi de probabilité d’apparition d’un événement, sans que cet événement soit prédictible, on se retrouve avec un système aléatoire. C’est le hasard, un événement dû à des causes indépendantes (par exemple la rencontre entre une tuile qui se détache d’un toit à cause du vent et la tête d’un passant : la tuile ne se détache pas parce que le passant passe, mais parce qu’il y a du vent : les évènements « la tuile se détache » et « le passant est là » ont des causes indépendantes). Là aussi, on pourrait sans doute prédire ce qui se passerait si l’on connaissait absolument tous les paramètres qui régissent le système, mais chaque événement a une certaine probabilité de se produire, ce qui n’est pas le cas du système chaotique.
Concepts basiques
Le hasard : un exemple pour mieux comprendre.
Examinons avec un exemple concret ce que signifie « tirage au hasard ».
Je vous donne
une pièce, vous la lancez en l’air. Si c’est face, vous la gardez, si
c’est pile, vous me la rendez. On recommence 1000 fois. Au bout du
compte, vous aurez gagné un certain nombre de pièces, aux alentours de
500 si elles ne sont pas truquées.
Le fait que vous
gagniez ou perdiez n’influe pas le fait que la pièce tombe sur pile ou
face : ce n’est pas parce que vous gagnez à face que les pièces vont se
mettre à miraculeusement tomber toutes sur face, ou toutes sur pile
(pour vous embêter). Il y a indépendance entre la face sur laquelle
tombe la pièce, et la face que vous devez obtenir pour gagner.
Il y a des
causes à la chute de la pièce : le mouvement que vous lui donnez, la
force avec laquelle vous la lancez etc. Si on connaissait précisément
tous les paramètres physiques qui gèrent la pièce, on pourrait prédire
sur quelle face elle va tomber, mais c’est indépendant du fait que vous
gagniez à face. Gagner à face n’influe pas le comportement de la pièce.
Donc, au final, est-ce que les pièces que vous avez dans votre main à la fin du jeu sont là « par hasard » ? Non.
Le hasard a joué
un rôle, mais la règle du jeu stipulant que vous aviez le droit de
gagner les pièces tombant sur face a aussi joué. Si cette règle avait
été différente, vous auriez eu, pour les mêmes tirages, un nombre de
pièces différent dans la main (on aurait pu dire que vous gagniez à
pile, ou alors que vous ne gagniez que dans le cas de deux piles
consécutifs etc...)
De la même
manière, en refaisant un essai de 1000 tirages, vous n’aurez
probablement pas le même nombre de pièces au final, même avec des
règles identiques.
En ce qui concerne l’évolution, le hasard joue un rôle au niveau de la dérive génétique, car il y a
toujours des allèles qui sont éliminés par hasard au cours du passage
d’une génération à l’autre (vous ne passez que certains de vos
caractères à vos enfants, les autres sont perdus). Il joue également au
niveau des mutations, car il y a indépendance entre la nature de la
mutation et son effet. Ce n’est pas parce qu’une mutation serait
favorable qu’elle va apparaître (nous verrons ceci dans le chapitre suivant). D’un autre côté, la sélection
naturelle élimine les mutations entraînant de caractères défavorables à
la reproduction de l’individu. C’est la « règle du jeu » non hasardeuse.
Pour la dérive génétique, on peut calculer le pourcentage de chances qu’un allèle a d’être transmis à la génération suivante, ou de disparaître, sur l’ensemble de la population, sachant sa fréquence initiale. Par exemple, un allèle porté par un seul individu, et ne conférant à cet individu aucun avantage, ni désavantage, c’est à dire neutre dans ses effets sur la reproduction, a une chance sur deux de disparaître immédiatement à la génération suivante si cet individu n’a qu’un seul descendant – mais on ne peut pas prédire ce qui va se passer en réalité, on ne peut parler que de probabilités...
Et oui, puisque nous possédons deux allèles pour chaque gène, nous avons une chance sur deux de transmettre chacun des allèles en question à chaque descendant... Sachant que l’effectif théorique nécessaire au renouvellement d’une population est de deux descendants par femelle, vous pouvez compter les chances qu’a chaque allèle d’être transmis en moyenne...
D’ailleurs, attardons-nous un instant sur cet effectif théorique nécessaire au renouvellement d’une population. Imaginez une population de deux cents individus (des moutons par exemple), avec cent mâles et cent femelles. Pour que l’effectif de cette population reste fixe, sans diminuer, ni augmenter, il suffit que chaque femelle ait deux petits. Si l’on considère que les petits ont une chance sur deux d’être de chaque sexe, cela nous donnera, à la génération suivante, grosso modo 100 mâles et 100 femelles. Or, vous connaissez tous des espèces qui ont beaucoup plus de deux petits au cours de leur vie. Certains poissons en ont des centaines (et ne parlons pas des mouches). Si tous ces petits arrivaient à maturité, ils envahiraient purement et simplement la planète !
Comme cela n’est pas le cas, cela signifie que, sur la grande quantité de descendants produits, la plupart meurt avant d’être mature. Ils sont mangés par les prédateurs... ou n’arrivent pas à trouver assez de nourriture eux-mêmes, par exemple. Donc, chaque femelle produit en moyenne et en théorie, deux petits matures par génération...
Naturellement, certains individus arrivent à produire une descendance mature plus nombreuse que la moyenne des autres, que ce soit par chance, indépendamment des aptitudes que leurs confèrent leurs gènes, ou parce qu’ils possèdent un avantage génétique quelconque leur donnant un avantage sélectif. Les caractères qu’ils portent, s’ils sont héritables, ont donc de grandes chances de se répandre dans la population au fil des générations, comme nous l’avons vu plus haut, dans le chapitre sur la sélection.
Dans une population de petite taille, quand un allèle est rare, il peut n’être porté que par quelques individus. Que se passe-t-il alors si, pour une raison fortuite, ces individus ne peuvent se reproduire ? L’allèle disparaît, et ce même s’il était avantageux. Car être avantageux ne signifie pas « garantir » la reproduction. On peut porter un super allèle, et se prendre une branche sur la tête... Ou bien, dans le cas de la reproduction sexuée, on peut posséder cet allèle en un seul exemplaire (hétérozygotie*), et, par hasard, transmettre l’autre allèle à ses descendants.
Pourtant, de très nombreux allèles ne sont ni avantageux ni désavantageux par rapport aux autres. On les appelle les allèles neutres. La sélection naturelle n’a pas de prise sur eux. Leur fréquence dans la population ne se modifie que grâce à la dérive génétique.
Regardez la figure 1.10 : elle représente une petite simulation (réalisée avec le logiciel R13). Imaginons dix gènes, chacun représenté par une couleur différente, qui ont chacun deux exemplaires (allèles). Chaque allèle est présent dans la moitié de la population, ils sont à une fréquence de 0,5. Ils ont à chaque génération une chance sur deux d’être transmis à chaque descendant. Il y a 100 individus dans la population, et chacun a un seul descendant. Que va-t-il se passer au bout de plusieurs générations ?
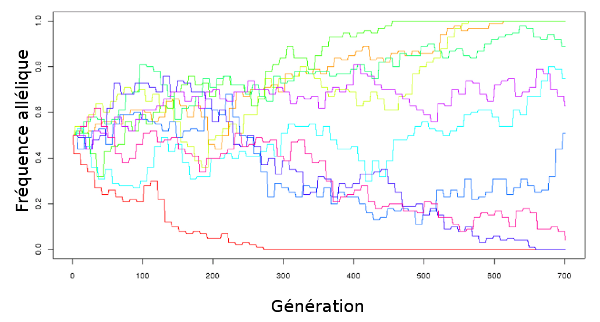 |
| Figure 1.10 Simulation de dérive génétique simple : on part de dix gènes à deux allèles, chaque allèle étant à la fréquence initiale de 0,5 puis on les laisse évoluer pendant 700 générations. |
Comme vous pouvez le voir, au bout de 700 générations, quelques allèles sont « fixés » (c’est à dire qu’ils ont atteint une fréquence de 1 ou de 0, donc ils sont seuls ou ont disparu). Mais beaucoup ne sont pas fixés.
Refaisons l’expérience, mais cette fois-ci, pour chaque gène, un allèle est davantage représenté que l’autre (la fréquence initiale n’est plus à 0,5). Le résultat est présenté à la figure 1.11 .
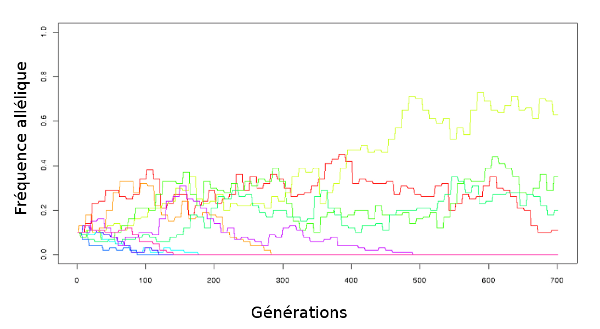 |
| Figure 1.11 Simulation de dérive génétique simple : on part de dix gènes à deux allèles, chaque allèle étant à la fréquence initiale de 0,1 puis on les laisse évoluer pendant 700 générations. |
Cette fois, beaucoup d’allèles ont disparu, mais certains ont quand même vu leur fréquence augmenter. En fait, la probabilité de fixation d’un allèle, c’est
à dire la probabilité que tous les individus de la population finissent
par porter cet allèle, est strictement égale à sa fréquence de départ. C’est à dire qu’en réalité, la probabilité de fixation d’un nouvel allèle est égale à la probabilité d’apparition de cet allèle (mutation, voir chapitre suivant).
Pour aller plus loin
Taux de fixation, taux de mutation, autre explication
Imaginez
une population au nombre fixe de deux individus, se reproduisant par
clonage. À chaque génération, il y aura deux nouveaux individus, qui remplaceront les anciens. Voyons
ce qui se passe à un point précis de leur génome, qui est par exemple
occupé par une base « A », pour « adénine ». À chaque génération, cette
base a une certaine probabilité d’être mutée, c’est à dire d’être
remplacée par un T, un C ou un G. Appelons ce taux de mutation m. Dans
la population, on aura donc 2*m chances d’observer une mutation.
En
outre, chaque mutation a une chance sur deux d’être fixée par la suite
(puisqu’il n’y a que deux individus en tout). Si la population était constituée de N individus au lieu de 2,
nous aurions N*m chances d’observer une mutation, et cette mutation
aurait 1/N chances d’être fixée. Le taux de fixation étant égal à la
probabilité d’observer une mutation multiplié par le taux de fixation,
cela nous donne :
Taux de fixation = $N*m*1/N$ = m = Taux de mutation
On pourrait s’amuser à faire comme ça des milliers de simulations, et même si on partait à chaque fois de la même situation initiale, on n’obtiendrait jamais deux fois le même résultat. C’est pour cela que les résultats sont « imprédictibles ».
Vous avez pu voir néanmoins que la dérive génétique nécessite pas mal de temps (700 générations, prennent par exemple environ 700 ans au minimum chez le chien, qui peut avoir des petits au bout d’un an). Et le temps nécessaire est plus long dans les populations de grande taille. Mais l’évolution se déroule sur des millions d’années...
La dérive génétique est une force évolutive très importante, car son rôle peut-être plus fort encore que celui de la sélection naturelle dans l’évolution d’une population.
c. En fin de compte, à quel niveau s’effectuent la sélection naturelle et la dérive génétique ?
Prenons un exemple, celui des lions. Vous savez peut-être que les lions mâles s’approprient une bande de lionnes, avec lesquelles ils vivent jusqu’à être évincés par un autre lion, plus jeune, plus fort. Or, quand un lion prend possession d’une nouvelle troupe, la première chose qu’il fait est de tuer les lionceaux. Pourquoi ?
Cela semble tout à fait idiot pour la « survie de l’espèce ». Tous ces lionceaux avaient déjà un certain âge, il avait été coûteux de les produire, de les allaiter, et voilà tout réduit à néant par un lion stupide.
 |
Mais en fait... que se passe-t-il une fois les lionceaux tués ? Les lionnes, qui n’ont plus personne à allaiter, retombent bientôt en chaleurs. Et le lion peut les féconder.
Imaginez deux lions. Le lion Marcel possède un allèle qui détermine un comportement « tueur de lionceaux » (et oui, ce comportement est instinctif, c’est à dire génétiquement déterminé). Le lion Jean ne le possède pas. Chacun prend possession d’une troupe de lionnes pendant quelques années, avant d’être évincé à son tour. À son arrivée, Marcel tue les lionceaux et les lionnes sont en chaleurs. Quelques mois plus tard, naît une nouvelle troupe de lionceaux, portant à leur tour l’allèle « tueur de lionceaux ». Jean, lui, doit attendre un an avant d’avoir ses premiers lionceaux, le temps que les lionnes aient élevé les précédents. Qui au final aura eu le plus de lionceaux ? Le tueur de lionceaux. Ce comportement est observé chez un certain nombre d’espèces, dont des primates14.
Nous avons vu précédemment que, du point de vue de la sélection, peu importe qu’un comportement ou un caractère physique soit défavorable à l’espèce, s’il est favorable à la transmission des gènes qui codent pour lui. Il peut même entraîner la mort précoce de l’individu ; si cette mort ne nuit pas à la production d’une descendance mature au moins aussi nombreuse que celles des autres membres de l’espèce, le caractère ne sera pas contre-sélectionné... Cela explique pourquoi, même si les grands-parents peuvent parfois aider substantiellement à élever les enfants, tant de maladies nous atteignent plus particulièrement pendant notre vieillesse : cancer, alzheimer etc... C’est simplement qu’elles n’ont jamais été contre-sélectionnées — et pour cause, de toutes manière nos ancêtres ne vivaient pas à un âge assez avancé pour en être victimes... Peut-être que si nous pouvions vivre 200 ans, des maladies de vieillesse encore plus graves, dont nous n’avons pas idée, s’abattraient sur nous...
Réfléchissons maintenant à la reproduction. Nous avons coutume de dire que les individus « se reproduisent ». Est-ce exact ? Non, car nos descendants sont différents de nous-même. Même si ils portaient le même génome que nous, s’ils étaient nos clones, comme dans le cas de la reproduction asexuée que pratiquent certaines espèces, ils ne seraient pas « nous ». Que leur avons nous transmis ? Nos gènes. La moitié de notre information génétique, transmise par un ovule ou un spermatozoïde, dans le cas des espèces sexuées.
Pour aller plus loin
L’information portée par les gènes... subtilité
Voyons comment se transmet l’information portée par les gènes... Chez les bactéries, l’ADN n’est
pas enfermé dans un noyau, et les chromosomes sont circulaires. Elles
possèdent en plus des fragments plus petits d’ADN, nommés plasmides,
qu’elles peuvent échanger. Ceci est une manière de pratiquer le sexe, c’est à
dire l’échange d’information génétique, sans se reproduire !
Chez
les eucaryotes, c’est à dire ceux dont l’ADN est enfermé dans un noyau,
comme les plantes, les champignons et les animaux, la situation est
aussi un poil compliquée.
 |
En
effet, les plantes, comme les animaux, possèdent dans chacune de leurs
cellules des « organites » (les « organes » de la cellule) dont l’origine
est parfois particulière. Ainsi que l’a découvert Lynn Margulis15, certains sont en effet des bactéries, qui
vivent en symbiose avec les organismes depuis des millions d’année, et
qui leurs rendent divers services, en échange du gîte et du couvert. Cependant, tous les organites ne sont pas issus de bactéries symbiotiques,
seulement quelques-uns.
Par
exemple, il y a les les mitochondries, qui permettent à la cellule d’utiliser l’oxygène
pour fabriquer de l’énergie. Chez les plantes, on trouve aussi les chloroplastes, qui
contiennent la chlorophylle et permettent de transformer l’énergie
lumineuse reçue par la plante en sucre.
Ces organites particuliers ont leur propre ADN.
Chez
les animaux, ou les végétaux, ces organites passent d’une génération à l’autre presque
exclusivement par les ovules, qui en contiennent plein, alors
que le spermatozoïde ne donnera que son noyau.
Vous
avez donc reçu un petit plus d’information génétique de votre mère que
de votre père ; sans parler des éventuels « effets maternels », vus en 1.1 – les « effets paternels » sont beaucoup plus rares.
La seule chose qui « passe » d’une génération à l’autre, c’est donc de l’information génétique (et épigénétique, voir encadré1.1 ). C’est donc cette information qui est la cible de la sélection et de la dérive (ni l’espèce, ni l’individu... qui est si éphémère).
Selon ce concept, les individus se retrouvent assimilés à des « avatars » que leurs gènes ont créés pour assurer leur reproduction16. Les individus ne sont pas la cible directe de la sélection, notamment car, d’une génération à l’autre, les combinaisons d’allèles changent au fur et à mesure des croisements successifs. Il y a aussi une possibilité d’apparition de nouveaux allèles par mutation (voir chapitre suivant). Par conséquent, chaque allèle peut être considéré individuellement.
Néanmoins, le rôle de l’individu est important, en tant que « présenteur » de gènes : la sélection ne peut toucher les allèles qu’au travers des combinaisons alléliques des individus. Ainsi, un individu peut porter un allèle très favorable dans ses conditions environnementales propres, il ne sera pas transmis s’il porte en même temps un autre allèle, qui entraîne, par exemple, la stérilité.
Cette idée de la sélection des gènes au détriment des individus est nommée « théorie du gène égoïste » et a été développée, entre autres, par Richard Dawkins. Selon cette idée, n’existent à l’heure actuelle que les gènes ou allèles qui se sont le mieux reproduits, au détriment des autres gènes. Les gènes qui se reproduisent le mieux sont ceux qui produisent les organismes les plus aptes à les reproduire.
Cette théorie, que certains trouvent « choquante », a permis de résoudre beaucoup de questions qui semblaient insolubles, notamment au niveau de l’étude des comportements des animaux, comme l’infanticide chez les lions. Elle permet aussi d’expliquer les bases de la coopération chez les fourmis.
Comme les abeilles et les guêpes, les fourmis peuvent former, selon les espèces, des colonies de quelques dizaines (parfois moins) à plusieurs centaines d’individus. Dans une colonie, on trouve une reine, et les ouvrières, ses filles. Les ouvrières sont toujours des femelles, nous allons voir pourquoi. Enfin, la colonie compte potentiellement quelques mâles et quelques futures reines.
Les fourmis partagent avec leurs cousines guêpes et abeilles une particularité très spéciale. Seules les femelles portent deux exemplaires de chaque chromosome, et donc sont diploïdes* (voir figure 1.4 pour la différence entre haploïde et diploïde). Les mâles sont haploïdes, c’est à dire qu’ils n’ont qu’un seul lot de chromosomes. Lorsque la reine pond un œuf, elle a le choix : fécondé d’un spermatozoïde, cet œuf donnera une femelle, et non fécondé, il donnera un mâle.
La plupart des femelles sont stériles : ce sont les ouvrières. La question est : pourquoi les gènes des fourmis peuvent-ils les autoriser à renoncer à la reproduction ?
C’est William Hamilton qui, se basant notamment sur les travaux de Fisher17 et Haldane18, a eu une idée de génie19 : il a calculé le degré d’apparentement des ouvrières (toutes sœurs) entre elles, avec la reine, et avec leur potentielle descendance. Et là, surprise ! Si les ouvrières ont le même père, elles sont davantage apparentées à leurs sœurs qu’à une hypothétique descendance : elles ont plus de gènes en commun.
Par conséquent, quand une ouvrière prend soin de sa sœur, ou de la reine, elle favorise davantage la transmission de ses propres gènes que si elle avait elle-même des petits. D’autres mécanismes peuvent encore renforcer l’altruisme des ouvrières20, en particulier si elles sont demi-sœurs, mais une base fondamentale est posée.
Autre exemple : un gène vraiment très égoïste chez certaines espèces de guêpes parasitoïdes21. Le terme « parasitoïde » indique que l’hôte est tué au cours du développement du parasite. Il s’agit par exemple des guêpes qui pondent leurs œufs dans des chenilles, les larves se nourrissant des tissus de la chenille, ce qui a pour conséquence de la tuer. Les guêpes ont un mode de reproduction semblable à celui de leurs cousines les fourmis. La femelle, après s’être accouplée, peut féconder un ovule avant de le pondre, ce qui donnera une femelle, ou ne pas le féconder, ce qui donnera un mâle. Or, certains mâles portent un chromosome très spécial, surnuméraire. Celui-ci a la propriété étonnante d’entraîner la perte de tous les autres chromosomes d’origine paternelle présents dans l’œuf, au début de son développement.
Si une reine est fécondée avec des spermatozoïdes portant un tel chromosome, imaginez ce qui arrive quand elle les utilise... L'embryon, au
lieu d'avoir deux lots de chromosomes par cellule, n’en possède qu’un,
issu de la mère, plus le chromosome surnuméraire, issu du père. Comme il
n’y a qu’un lot de chromosomes, l'embryon donne une guêpe mâle, comme si
l’ovule n’avait jamais été fécondé. Mâle qui va, à son tour, répandre ce fameux chromosome surnuméraire !
Cela nous amène donc à la question : une espèce peut-elle s’éteindre à cause de quelques « gènes égoïstes » ? C’est facile à imaginer avec l’exemple précédent, mais on peut aussi imaginer un autre exemple : il suffirait d’un allèle favorisant la monopolisation des ressources (nourriture, espace...) par son porteur, mais ne lui permettant pas de produire beaucoup de descendants. Privant les autres individus de ressources, il les priverait de reproduction, et donc se répandrait dans la population, diminuant le nombre de descendants fertiles d’une génération à l’autre... et donc rendant l’espèce sensible à l’extinction22. On est loin de l’idée de la « sélection pour la survie du plus apte » ou la « survie de l’espèce ».
Cette idée du gène « égoïste », peut sembler troublante, et pose pas mal de questions philosophiques, sur lesquelles nous reviendrons. Mais en fin de compte, savoir que nos gènes nous ont développés « pour » les reproduire ne nous oblige pas à le faire...
Pour aller plus loin
Transmission verticale - transmission horizontale, et sélection
Nous venons de voir que la sélection s’effectue au niveau des gènes. Est ce que d’autres types d’information sont également susceptibles d’être sélectionnés, comme l’information épigénétique (voir 1.1 ) ? Pour que de l’information soit sélectionnée, il faut qu’elle soit transmise d’une génération à l’autre, or l’information épigénétique disparaît potentiellement en une ou deux générations. Néanmoins, si un caractère dû à un facteur épigénétique est défavorable pour l’individu qui le porte, il nuira à la reproduction de celui-ci en raccourcissant sa durée de vie, par exemple, ou pour une autre raison. Il nuira donc à sa propre transmission. Inversement, si les mécanismes épigénétiques apportent un avantage reproductif aux organismes qui les possèdent par rapport aux autres, la capacité à employer ces mécanismes sera sélectionnée.
Nous restons ici au niveau des transmissions dites « verticales », c’est à dire au niveau des caractères transmis d’une génération à l’autre par filiation. Néanmoins, il existe un autre mode de transfert, dit « horizontal », c’est à dire entre deux individus potentiellement de la même génération, et non nécessairement apparentés (fig 1.12 ). La maladie de la vache folle donne un exemple d’information épigénétique transmise horizontalement. En effet, cette maladie est dûe à une protéine nommée prion, qui existe en deux formes. Une forme dite « infectieuse » a la propriété de transformer la forme « normale » en « infectieuse ». Elle se propage ainsi dans la cellule (le neurone), et conduit à sa mort, puis s’infiltre dans les neurones voisins. Elle peut également passer d’un individu à l’autre par consommation de tissus contaminés (causant la propagation de la maladie de Creutzfel-Jacob, et
celle de l'encéphalite spongiforme bovine par exemple). Ainsi la transmission horizontale permet à la protéine de se propager dans la population. La reproduction de la protéine nuit à celle de l’individu, mais elle se propage quand même car la mort de l’individu n’est pas un obstacle à cette propagation, si les vivants consomment les tissus nerveux des morts.
| Figure 1.12 Transfert d’information horizontal (d’un individu à l’autre, qui ne descendent pas l’un de l’autre) et vertical (par descendance). |
L’information génétique peut aussi, chez certaines espèces, être transférée horizontalement. C’est un jeu très apprécié des bactéries, qui utilisent ainsi les avantages du sexe (le brassage génétique), sans passer par la case « reproduction ».
Chez l’homme, ainsi que chez d’autres espèces, notamment des grands singes, certaines espèces d’oiseaux etc., il existe un autre type de caractère transmis horizontalement. Il s’agit des comportements appris par apprentissage social (imitation etc.). Ces comportements sont également potentiellement soumis à la sélection naturelle. Ainsi, on peut imaginer que les différentes cultures culinaires humaines ont été sélectionnées en fonction de leur bonne adéquation avec les besoins physiologiques des humains. Ainsi, des chercheurs ont observé que les cuisines les plus épicées correspondaient aux pays les plus chauds, l’effet bactéricide des épices limitant la croissance des bactéries sur la nourriture, et en particulier la viande23. De manière générale, si une mode culinaire est inadaptée aux besoins humains, la nourriture ne permettra un bon développement des individus. On peut donc imaginer que l’ensemble de nos règles de cuisine ont été construites grâce à la combinaison de facteurs génétiques et culturels. Génétiques car nous avons un goût instinctif pour certains aliments (ce qui est bon au goût est souvent comestible, mais ce critère, même s’il nous permet d’éviter certains poisons ou aliment impropres à la consommation, n’est pas suffisant). Culturels car la reconnaissance et le mode de préparation des aliments est variable entre les populations et s’apprend socialement.
Mais quoi qu’il en soit, au final, ce qui est sélectionné est de nature informative.
d. Les flux de gènes
De la même façon que les éleveurs peuvent croiser différentes races de chiens pour obtenir des phénotypes* encore plus conformes à leurs souhaits, dans la nature les flux migratoires entre des populations d’une même espèce peuvent maintenir la diversité génétique et s’opposer à la réduction du nombre d’allèles par la sélection et la dérive. Le brassage génétique (voir encadré 1.3 ) qui résulte de la présence de nombreux allèles pour chaque gène permet ainsi de multiplier les combinaisons alléliques dans la population. Ainsi, les possibilités d’adaptation de la population augmentent.
Concepts basiques
Le brassage génétique - le sexe
Nous avons vu que dans le cadre de la reproduction sexuée, chaque parent transmet la moitié de son information génétique à sa descendance, la moitié de ses chromosomes.
Une partie du brassage s’effectue à ce niveau là : ce ne sont pas toujours les mêmes chromosomes qui se retrouvent dans tous les gamètes. Il y a autant de combinaisons que le permet le nombre de chromosomes (par exemple, si une espèce a 6 chromosomes, il y a 8 gamètes possibles pour chaque individu).
Une autre partie du brassage s’effectue par recombinaison, grâce aux « crossing overs ». Ce mécanisme permet l’échange de deux bouts de chromosomes au moment de la division méiotique (celle qui réduit le stock de chromosomes par deux pour fabriquer les gamètes). Deux chromosomes se croisent physiquement dans la cellule et échangent un bras, ou une partie de bras.
Grâce à ces deux mécanismes, pratiquement chaque gamète produit par un individu est unique. C’est pour cela que deux parents ne peuvent pas avoir deux enfants identiques à moins d’avoir des jumeaux monozygotes (une cellule-œuf se divise en deux cellules qui au lieu de rester ensembles, se séparent et donnent chacune un embryon, au bagage génétique strictement identique).
Le brassage génétique, c’est donc la conséquence principale du sexe. Il peut être intéressant, au passage, de noter que certaines espèces sont divisées en plus de deux sexes (un sexe étant un type d’individus n’ayant pas, biologiquement, la possibilité d’obtenir de descendance avec un autre individu du même type et de la même espèce). En outre, d’autres espèces sont divisées en deux sexes, mais pas de mâle ni de femelle. On ne parle de mâle et femelle que quand les gamètes sont de taille différente (le petit est le mâle, le gros la femelle). Si les deux gamètes sont de même taille, comme chez certaines levures, on nomme les sexes différemment (plus et moins, par exemple).
Chez les bactéries, le brassage génétique ne s’effectue pas au moment de la reproduction (division cellulaire), mais à d’autres périodes. Les bactéries échangent entre elles du matériel génétique par divers mécanismes, dont la conjugaison (simple transmission entre individus). Comme quoi, on peut être un organisme asexué, et pratiquer le sexe quand même...
1. source : wikipedia.
2. Imesch PD, Wallow IH, Albert DM. The color of the human eye : a review
of morphologic correlates and of some conditions that affect iridial
pigmentation. Surv Ophthalmol. 1997 Feb ; 41 Suppl 2 : S117-23. Review.
3. Sturm RA, Frudakis TN. Eye color : portals into pigmentation genes and ancestry. Trends Genet. 2004 Aug ; 20(8) : 327-32 (Nom des auteurs. Titre de l’article. Nom de la revue. Date de publication ; numéro de la revue (numéro du volume) : pages).
4. Bertina RM, Koeleman BP, Koster T, et al. Mutation in blood coagulation factor V associated with resistance to activated protein C. Nature. 1994 ; 369 (6475) : 64–7.
5. Adamzik M, Frey UH, Riemann K, Sixt S, Lehmann N, Siffert W, Peters J. Factor V Leiden mutation is associated with improved 30-day survival in patients with acute respiratory distress syndrome. Crit Care Med. 2008 Jun ; 36(6) : 1776-9.
6. Burger JM, Kolss M, Pont J, Kawecki TJ. Learning ability and longevity : a symmetrical evolutionary trade-off in Drosophila. Evolution. 2008 Jun ; 62(6) : 1294-304.
7. Buchholtz EA, Booth AC, Webbink KE. Vertebral anatomy in the Florida manatee, Trichechus manatus latirostris : a developmental and evolutionary analysis. Anat Rec (Hoboken). 2007 Jun ; 290(6) : 624-37.
8. Wetzel R. M. 1985. The identification and distribution of recent Xenarthra (Edentata). In The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas. Montgomery G. G. (ed.) pp 5–21 Smithsonian Institution Press. Washington, DC.
9. Borghans JA, Beltman JB, De Boer RJ. MHC polymorphism under host-pathogen coevolution. Immunogenetics. 2004 Feb ; 55(11) : 732-9. Epub 2004 Jan 13.
10. Zahavi, A. Mate selection - a selection for a handicap. Journal of Theoretical Biology 1975 ; 53 : 205-214.
11. Fisher, RA (1930) The Genetical Theory of Natural Selection, Oxford University Press, ISBN 0-19-850440-3.
12. Candolin U. The relationship between signal quality and physical condition : is sexual signalling honest in the three-spined stickleback ? Anim Behav. 1999 Dec ; 58(6) : 1261-1267.
13. R Development Core Team. R : A language and environment for statistical computing. 2004. ISBN 3-900051-00-3, site internet : http://www.R-project.org.
14. Hrdy SB. Infanticide as a primate reproductive strategy. Am Sci. 1977 Jan-Feb ; 65(1) : 40-9.
15. Sagan L. On the origin of mitosing cells. J Theor Biol. 1967 Mar ; 14(3) : 255-74.
16. cf « Les avatars du gène », P.H. Gouyon.
17. Fisher, R. A. (1930). The Genetical Theory of Natural Selection, ISBN 0-19850440-3.
18. Haldane, J. B. S. Population Genetics. New Biology. 1955) ; 18 : 34–51.
19. Hamilton WD. The genetical evolution of social behaviour.J Theor Biol. 1964 Jul ; 7(1) : 1-52.
20. Wenseleers T, Ratnieks FL. Enforced altruism in insect societies. Nature. 2006 Nov 2 ; 444(7115) : 50.
21. Werren JH, Stouthamer R. PSR (paternal sex ratio) chromosomes : the ultimate selfish genetic elements. Genetica. 2003 Jan ;117(1) :85-101. Review.
22. Rankin DJ, López-Sepulcre A, Foster KR, Kokko H. Species-level selection reduces selfishness through competitive exclusion. J Evol Biol. 2007 Jul ; 20(4) : 1459-68.
23. Sherman PW, Hash GA. Why vegetable recipes are not very spicy. Sherman PW, Hash GA. Evol Hum Behav. 2001 May ; 22(3) : 147-163.